
Vitamin K may not be as popular as vitamin C, but it sure is important! In fact, research in the recent years has shown that there are vitamin K concentrations in the brain and receptors which react to the presence of vitamin K. Furthermore, given its antioxidant nature, vitamin K may be able to act as a neuroprotectant, shielding the brain from oxidative stress and the diseases which may develop as a result of stress. Given the limited knowledge on vitamin K, more research is necessary, in order to uncover its nature and mechanisms at work.
In this article, we will examine what vitamin K does in the brain and review recent studies and animal models which explore the relationship between vitamin K and behavior.
Vitamin K and the Brain
Vitamin K, as many of the other vitamins, comes in many shapes and sizes. Some forms of vitamin K are
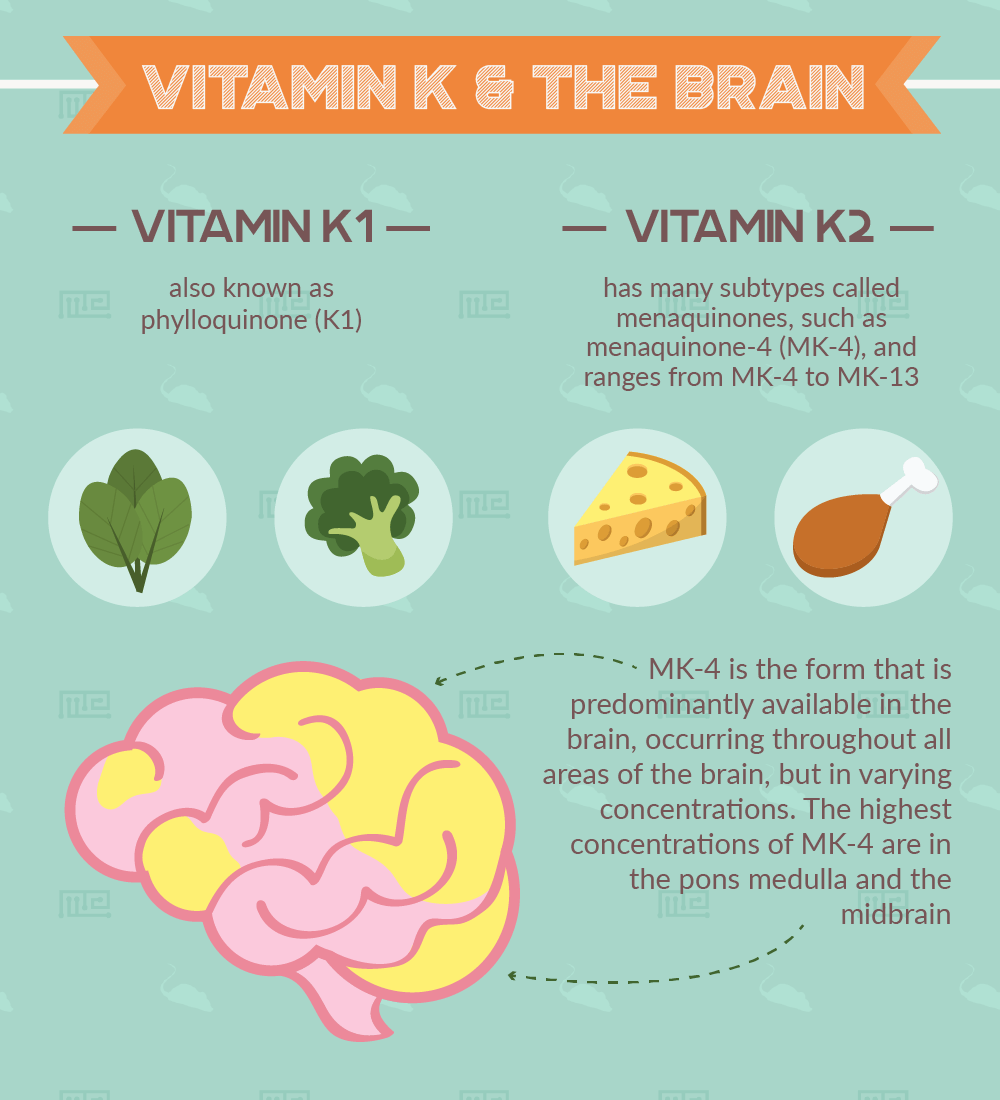
- Vitamin K1 also known as phylloquinone (K1)
- Vitamin K2, its subtypes are called menaquinones, such as menaquinone-4 (MK-4)
MK-4 is the form that is predominantly available in the brain, occurring throughout all areas of the brain, but in varying concentrations. The highest concentrations of MK-4 are in the pons medulla and the midbrain [1].
The availability and distribution of vitamin K as form MK-4 in the brain depends on several factors, including nutrition. Gender is also another variable of concern. For example, female Brown Norway rats have been shown to have higher MK-4 concentrations in the cerebellum and cortex than male rats do [2].
Research, however, which links vitamin K to behavior and cognition remains extremely limited.
Vitamin K and Other Brain Molecules
Vitamin K and Sphingolipids
Correlations have been observed between vitamin K and sphingolipids, a class of complex lipids which can be found in any mammalian cell acting as crucial components of cellular membranes. Sphingolipids are very important to the central and peripheral nervous systems wherein they can be found in quite high concentrations. The fact that there is a correlation between vitamin K and such a vital lipid for the central nervous system is noteworthy for future research.
The major sphingolipids are sphingomyelin, ganglioside, cerebroside, ceramide, and sulfatide. They occur in various concentrations and proportions, exhibiting complex relationships amongst each other which still remain to be fully grasped. For example, as will be further discussed in the upcoming sections, some sphingolipids may be low or high in one condition and then exhibit a completely different profile in another condition.
Sphingolipids are very important constituents in brain physiology with multiple roles and functions [3]. They are crucial for maintaining the cell’s integrity by providing a stable cellular membrane. Also, sphingolipids are believed to have cell signaling capabilities.
Vitamin K-dependent Proteins
Another way that Vitamin K impacts the brain is through vitamin K-dependent proteins such as Gas6[4] and Protein S. Both of these proteins are mediated by vitamin K and are a part of brain physiology, but have yet to be explored deeply by behavioral research.
Gas6, discovered in 1993 and named so due to its relationship with the growth arrest-specific gene 6, is expressed in several brain regions including the hippocampus (the dentate gyrus, and areas CA1 and CA3), the thalamus, hypothalamus, cerebral cortex, midbrain, and cerebellum (particularly the Purkinje neurons) [5]. Although the Gas6 protein is present in the brain, there is an extremely limited body of research pertaining to the link between behavior and this vitamin K-dependent protein.
Protein S, in the adult nervous system, has been observed in the choroid plexus, the locus coeruleus, and astrocytes. However, animal studies, which are more invasive in nature, have demonstrated that protein S mRNA can also exist in dentate gyrus’ granule neurons and in pyramidal neurons of the hippocampus and cortex. In contrast to the Gas6 protein, the research available on protein S is even smaller in scope, but it is believed that this protein is involved somehow in neuroprotection [6].
Aging, Sphingolipid Metabolism, and Vitamin K
As mentioned previously sphingolipids are integral to the brain’s physiology and vitamin K has been demonstrated to correlate with sphingolipids.
In a recent study with geriatric patients, vitamin K intake was observed and subsequent behavior profiles noted [7]. The geriatric participants which had low vitamin K intake scored significantly lower in the Mini Mental State Exam, a cognitive tool for assessing mental function, than those with medium to high vitamin K intake. In a different study, seniors with high serum concentrations of phylloquinone had higher test scores in their verbal episodic memory performances than those with low levels [8], suggesting that the relationship between vitamin K and cognition can affect information processing systems such as memory.
All of the major sphingolipids (sphingomyelin, gangliosides, cerebrosides, and sulfatides) have been demonstrated to change in both animals and humans as a result of aging [3]. Due to the interaction between sphingolipids, age, and the brain, as well as the correlation between vitamin K and sphingolipids, this is a potentially fruitful area for future research.
Sphingolipids are affected by aging. Aging changes sphingolipid metabolism which in turns affects brain physiology and, ultimately, behavior and cognition.
Manipulating Sphingolipids Causes Behavioral Alterations
In an interest to see whether manipulating ceramides, a particular type of sphingolipid, would have any effect on behavior, a group of researchers designed an experiment for that purpose. The experiment involved blocking an enzyme, neutral sphingomyelinase (nSMase2), crucial for ceramide synthesis, then they would measure the animals’ behavioral profiles. Their line of reasoning stems from the fact that the neurons of the dentate gyrus and the hippocampus are remarkably high in nSMase2 concentrations, implying that this enzyme is quite important for hippocampal-related memory formation [9].
So, the researchers set out to use inhibitor GW4869 via intraperitoneal injections, a method that is able to block nSMase2 activity in animal models. The control group received saline in their intraperitoneal injections. The administration began 7 days prior to testing and lasted for an additional two weeks, making a total of 21 injections.
After creating a control and an experimental group, the researchers measured the animals’ behavior, in order to determine whether manipulating nSMase2 would have any impact on behavior and cognition.
The researchers expected to see changes in memory and learning, given the fact that nSMase2 is abundant in brain regions known to be important for learning and memory.
To test spatial learning and memory abilities, a Morris Water Maze was used. The test was given 7 days after the GS4869 treatment began. Both experimental and control groups had equal swimming speed. However, when it came to learning the location of the platform through repeated trials, the experimental group did not show a decrease in latencies as the control group did. Furthermore, the analysis showed that the GS4869 mice spent less time in the target quadrant and in the area that surrounds the platform. The researchers interpreted these behavioral findings to mean that the mice with nSMase2 inhibited had a delay in forming spatial reference memories [9].
In the Radial Arm Water Maze, a maze with more spatial complexity than the Morris Water Maze that is used to measure episodic-like working memory, the control mice again outperformed those receiving the GW4869 inhibitor, indicating further impairments associated with the blocking of ceramide synthesis. In the Radial Arm Water Maze, the location of the platform changes on a daily basis, so each time the mice have to learn a new location. The GW4869 group had increased latencies in locating the platform in the subsequent trials and they also had fewer correct arm entries than the control group, indicating they were performing at chance level. Due to these additional behavioral findings, the researchers also concluded that nSMase2 inhibition (and thus the interference with ceramide synthesis) is also associate with issues in episodic-like working memory.
Some Vitamin K-Deficiency-Related Cognitive Problems
Inadequate vitamin K levels can lead to cognitive problems if the vitamin K deficiency has persisted over a long period of time.
One study, comparing the different behavioral responses of vitamin K deficiency over time, found differences in rats’ learning ability over time. The researchers employed a longitudinal study and tested the rats at specific points in time. The experiment began with 5-month-old-rats when the dietary inductions began. The rats were divided into three groups, receiving diets with low, adequate, or high vitamin K. Certain rats were subjected to behavioral tests at 6 months of age, then at 12 months of age, and then, finally, at 20 months of age, in order for the researchers to establish how prolonged vitamin K deficiency takes its toll on cognition and behavior compared to adequate and high intake levels.
The rats that were vitamin K deficient at 6 and 12 months of age did not show any learning problems, only at 20 months old did learning difficulties start to surface. The 20-month-old rats had the worst performance in the Morris Water Maze out of all the groups. These rats had the longest latency to find the hidden platform, meaning it took them the most amount of time to reach the platform [10]. However, during the cue test and the probe trial, statistical analysis showed that the rats’ performance remained unaffected by diet.
Then, at various points during the experiment, the researchers would sacrifice the animals in order to analyze the rats’ brain levels of vitamin K and sphingolipids at a certain stage in the experiment. The rats receiving high vitamin K diets had 7-times higher MK-4 concentrations than the rats in the vitamin K-deficient group. In the 20-month-old rats which were on high supplementation of vitamin K, they exhibited the lowest concentrations of ceramides (a type of sphingolipid) in the hippocampus than the other two conditions. Also, compared across diets, there were lower concentrations of cerebroside and sulfatide in the pons medulla in the high vitamin K diet than in the low and adequate conditions, but there were high concentrations of ganglioside in the midbrain and the pons medulla in the high vitamin K condition than the other two [10].
Given the behavioral findings, that vitamin K deficiency affected latency times but not other measurements (such as the probe trial), further research is required to investigate the nature of such observations.
Vitamin K Deficiency Decreases Locomotor Activity
Locomotor activity is an integral behavior in daily life and can be affected by vitamin K deficiency.
One group of researchers set out to study the effects of vitamin K deficiency on locomotor activity by inducing vitamin K deficiency in rats either through dietary depletion or warfarin treatment and then subjecting the rats to behavioral testing.
The vitamin K deficient rats, the ones that were induced with warfarin, demonstrated a general reduction of exploratory behaviors in the Open Field Test. To reach such a conclusion, the researchers used the composite measure of exploratory behaviors (rearing + walking + walking through center square) and compared it with the other groups’ composite score. Then, the researchers also compared the composite measure of less exploratory behaviors (freezing + sniffing + grooming + miscellaneous such as licking the apparatus) across groups. The warfarin vitamin K deficient rats had significantly reduced exploratory behaviors and increased the frequency of the less exploratory behaviors.
On the other hand, the dietary vitamin K-deficient rats showed decreased locomotor activity in the Radial-Arm Maze. The vitamin K-deficient rats performed significantly fewer choices than the control animals, demonstrating a decrease in overall movement and locomotor activity. However, the vitamin K-deficient rats were still able to perform the task correctly, they just were not as active and energetic as the controls. When the researchers reintroduced vitamin K to the dietary-depleted group of vitamin K, the rats did not demonstrate immediate improvement in their locomotor activity. It took three additional weeks of vitamin K supplementation to get the deficient rats back to baseline [11].
Although it has been noted that there is a connection between vitamin K and locomotor activity, the precise mechanisms by which vitamin K deficiency acts to inflict decreased locomotor activity remains unknown.
Conclusion
Future research tying vitamin K to cognition and behavior is absolutely necessary. Given the connection between vitamin K and sphingolipids and the abundance of sphingolipids in the brain, possible underlying relationships between biology and behavior could be uncovered.
Other areas for future investigation include the expansion of animal models. For example, different animal models may react differently to vitamin K deficiency, displaying nonidentical behavioral profiles as happens with vitamin D. Currently, warfarin models and methods that include enzyme-inhibition are popular, but more development is necessary, especially in the realms investigating the connection between vitamin K and memory.
References
- Carrié, Isabelle, et al. “Menaquinone-4 concentration is correlated with sphingolipid concentrations in rat brain.” The Journal of nutrition 134.1 (2004): 167-172.
- Huber, Agnes M., et al. “Tissue phylloquinone and menaquinones in rats are affected by age and gender.” The Journal of nutrition 129.5 (1999): 1039-1044.
- Denisova, Natalia A., and Sarah L. Booth. “Vitamin K and sphingolipid metabolism: evidence to date.” Nutrition reviews 63.4 (2005): 111-121.
- Manfioletti, G., et al. “The protein encoded by a growth arrest-specific gene (gas6) is a new member of the vitamin K-dependent proteins related to protein S, a negative coregulator in the blood coagulation cascade.” Molecular and cellular biology 13.8 (1993): 4976-4985.
- Prieto, Anne L., et al. “Gas6, a ligand for the receptor protein-tyrosine kinase Tyro-3, is widely expressed in the central nervous system.” Brain research 816.2 (1999): 646-661.
- Ferland, Guylaine. “Vitamin K and the nervous system: an overview of its actions.” Advances in nutrition 3.2 (2012): 204-212.
- Chouet, Justine, et al. “Dietary vitamin K intake is associated with cognition and behaviour among geriatric patients: the CLIP study.” Nutrients 7.8 (2015): 6739-6750.
- Presse, Nancy, et al. “Vitamin K status and cognitive function in healthy older adults.” Neurobiology of aging 34.12 (2013): 2777-2783.
- Tabatadze, Nino, et al. “Inhibition of neutral sphingomyelinase‐2 perturbs brain sphingolipid balance and spatial memory in mice.” Journal of neuroscience research 88.13 (2010): 2940-2951.
- Carrié, Isabelle, et al. “Lifelong Low-Phylloquinone Intake Is Associated with Cognitive Impairments in Old Rats–3.” The Journal of nutrition 141.8 (2011): 1495-1501.
- Cocchetto, David M., et al. “Behavioral perturbations in the vitamin K-deficient rat.” Physiology & behavior 34.5 (1985): 727-734.