
Why is it that we are able to recall a childhood birthday that occurred years ago, but we fail to remember what we had for lunch last Tuesday? Why is it that some memories are fleeting, lasting only seconds, while others may persist indefinitely? The difference lies in the unique way the brain stores, maintains and retrieves these enduring multisensory representations of the world known as Long term memories.
In this article, we will overview Long term memory, including its neurobiological correlates and common behavioral assays used to measure its performance in rodents and humans. Then, we will discuss common diseases and drugs that are known to affect this memory system.
What is Long term memory?
Long term memories are those you can recall days, months, or even years after they were originally stored. Unlike short-term memory, which has a finite capacity of approximately four items, Long term memory has a limitless store, but limited accessibility.
Long term memory is broadly divided into explicit, also known as declarative memory, and implicit, or non-declarative memory. Explicit memories are simply memories of things we can talk about. This is further broken down into semantic memory, the memory of factual knowledge, and episodic memory, the memory of narratives and events.
On the other hand, implicit memory involves knowledge for how to do things that are automatic and unconscious. One of the most common forms of implicit memory is procedural memory. Procedural memories are automatically retrieved and used to allow us to perform integrated cognitive and motor skills, such as riding a bike, driving a car, or tying shoes.[1]
Long term memory and the brain
Molecular and cellular underpinnings of LTM
Since the late 19th century it has been known that memories are stored as activity-dependent changes in synaptic connectivity. Only relatively recently have neuroscientists uncovered the precise molecular mechanisms behind memory encoding and maintenance.
While short-term memory involves functional changes in synapses, Long term memories require new RNA and protein synthesis. That is to say, new gene transcription and protein synthesis occur in response to repeated signals at the synapse, which go on to create anatomical changes in synaptic structure. This could include the growth of new synaptic connections, dendritic spines, new glutamate receptors, and the enhanced release of glutamate.
The most important molecular players in the formation of enduring memories include CREB, or cAMP response element-binding protein. When a postsynaptic receptor is activated with a signal like serotonin, this increases a second messenger molecule called cyclic AMP intracellularly in the postsynaptic cell. Cyclic AMP goes on to activate a series of intracellular signal transduction cascades down at the nucleus. These cascades recruit protein kinases that activate the transcription factor CREB. CREB then binds to promoters within the nucleus and turns on effector genes that encode the proteins needed to strengthen the synapse or for the growth of new connections. This molecular mechanism is evolutionarily conserved across animals and forms the molecular basis of Long term memory formation.[2]
Protein synthesis also plays a crucial role in Long term memory maintenance, after initial memory storage. One protein that plays a major role in memory maintenance is CPEB3, a prion-like protein that is formed in the nucleus and brought down to synapses by another protein when memories are formed and recalled. When CPEB3 is knocked out in transgenic mice, new memories can be formed but not kept intact.[3][4]
Brain regions involved in Long term memory
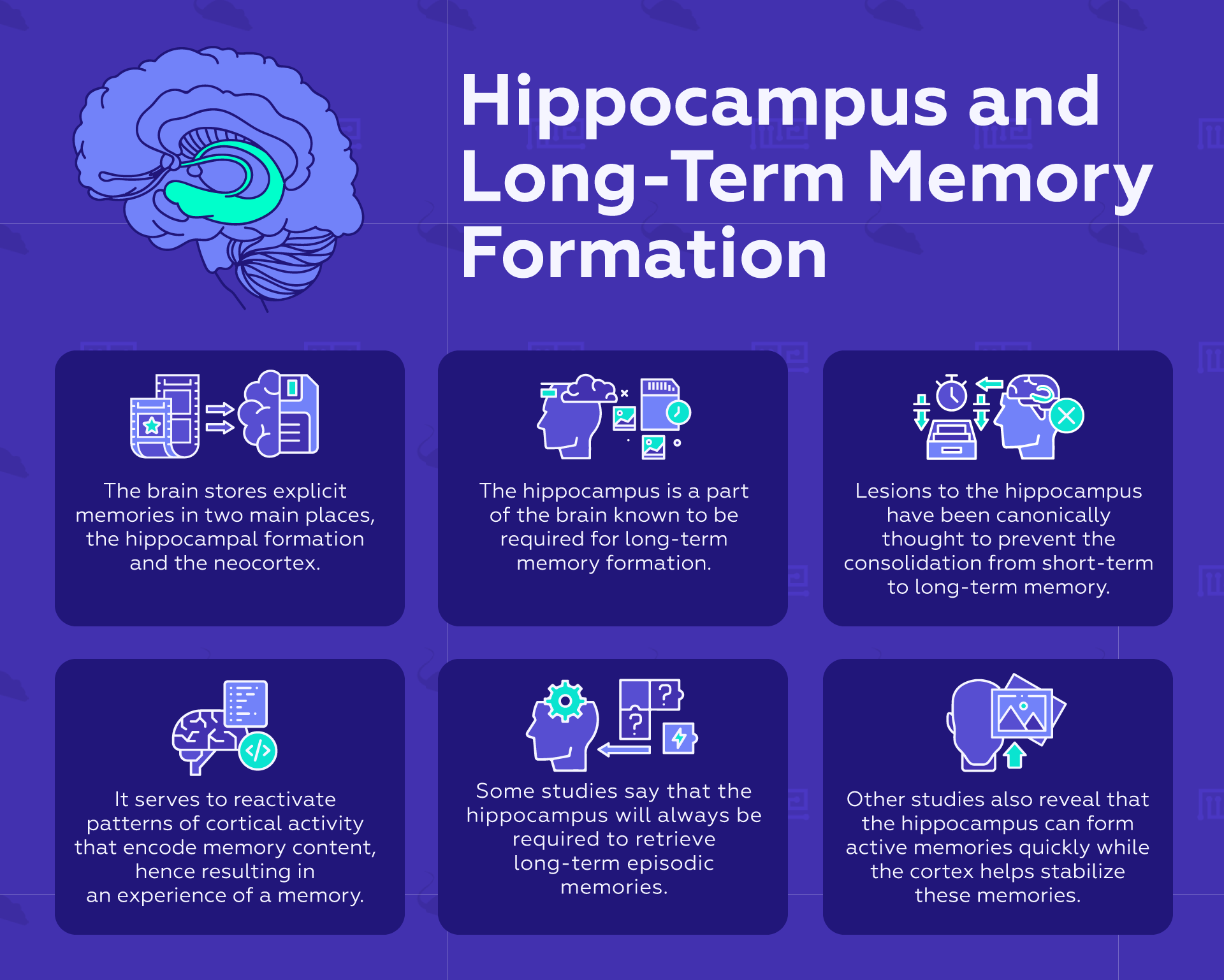
In general, memories are not stored within a designated memory center, but instead across a distributed network of brain regions. The brain stores explicit memories in two main places, the hippocampal formation, and the neocortex. Due to the automaticity of implicit memories, these memories are stored in the sensory cortices as well as the striatum, amygdala, basal ganglia, and cerebellum.[5]
The hippocampus is a part of the brain known to be required for Long term memory formation. Lesions to the hippocampus have been canonically thought to prevent the consolidation from short-term to Long term memory. These findings are derived from both animal experimental studies and human clinical studies. For the latter, the most famous case is patient H.M., whose hippocampus was removed to treat his epileptic seizures, thereby preventing his ability to form new memories. However, he could still retrieve some memories from before the surgery, which implies that Long term episodic memories may be stored without the involvement of the hippocampus.[6]
The indexing theory is a widely-accepted theory for how the hippocampus is involved in memory storage. In this theory, the hippocampus forms an index of cortical activity that was present during the actual experienced event, the contents of which are stored in distributed cortical networks. In this way, the hippocampal memory trace, when activated during memory retrieval, serves to reactivate the patterns of cortical activity that encoded the memory content, thereby resulting in an experience of a memory.[7]
The standard theory of consolidation maintains that cortical Long term memories are formed by a slow transfer from the hippocampus to cortical storage sites, the precise cortical region depending on the modality-specific information that is being processed. In this standard model of consolidation, the connections between cortical memory storage sites strengthen with time in a plasticity-dependent manner, so that hippocampal involvement in retrieval decreases with time. However, with conflicting research findings of hippocampal involvement in the retrieval of remote memories, the multiple trace theorists build on this theory and propose that the hippocampus would always be required to retrieve episodic memories, but not semantic memories. The latter are gist-like memories of abstract information, independent of the specific context in which they were formed.[7]
Tonegawa and colleagues at the Massachusetts Institute of Technology have recently proposed an alternative idea to the standard model of consolidation. In this theory, cortical engrams (memory traces) are formed at the same time as hippocampal memory traces, rather than a slow transfer over to the cortex from the hippocampus.[8]
These researchers labeled engrams, specific ensembles of cells associated with a particular memory, during a fear-conditioning event. They then could use optogenetic light to artificially reactivate the fear engrams to see if it could provoke a startle response from the mice.
What they found was that memory engrams for the fearful event were formed in both the hippocampus and prefrontal cortex. The difference is these cortical memory engrams are formed immediately at the time of learning in a “silent” state, at first inactivated by natural cues, but weeks later are unmasked fully. In their silent state, the memory could be activated and cause a startle response, but only artificially through optogenetic means. Two weeks later, Tonegawa and colleagues found that the cortical memory could be reactivated through natural cues, while the hippocampal engram fell silent, but could still be activated optogenetically. In this way, encoding is a simultaneous event but occurs strategically so that there is a division of labor: the hippocampus is able to form active memories quickly, whereas the cortex takes care of stabilizing the memory for the long term.[8]
Behavioral assays of Long term memory
In rodent studies, Long term memory is also referred to as reference memory. In the context of rodent experiments, this memory is defined as memory that remains constant over repeated trials. In this section, we will overview several common behavioral assays of Long term memory in rodents and humans.
Morris Water Maze
Developed by Richard Morris in 1981, the Morris water maze is one of the most widely used behavioral assays of Long term spatial learning and memory in rodents. It tests hippocampal-dependent learning, including the acquisition of Long term spatial memory. Long term memory performance is assessed during a probe trial, typically administered 1 day or more after the last learning trial. Multiple probe trials can be administered to determine the rate of memory consolidation.
Long term memory performance is indicated by behavioral measures such as platform-site crossovers, the gradual emergence of goal quadrant preference over days, escape latency, thigmotaxis duration, distance moved, and velocity during the time spent in the maze.[9]
Barnes Maze
The Barnes maze was developed by Carol Barnes in 1979 for use with rats (and later mice), with the advantage over the Morris water maze of not inducing stress in the animal with the requirement of swimming. Similar to the Morris water maze, the Barnes maze allows for the assessment of Long term spatial memory and learning. It is especially common in assessing spatial memory in Alzheimer’s disease models.
The maze’s protocol includes a habituation phase, a training phase, and a probe phase. Long term memory is assessed during the probe phase (typically a few days after the last training day), by removing the escape cage and observing the rodent’s behavior for a set amount of time. Mice with Long term memory intact will remember the location of the escape cage and will spend a greater-than-chance amount of time in this target quadrant.[10]
Fear Conditioning Paradigms
In fear conditoning paradigms, Long term memory is inferred from an increase in freezing behavior in rats in the presence of a conditioned stimulus (for instance, a light) that has been previously paired with an aversive unconditioned stimulus, such as a tail shock. The precision of this fear memory can be evaluated by comparing the freezing response in the fearful context to an alternate context (with no shock) across multiple delays in a context discrimination procedure. Hippocampal lesions that occur disrupt conditioned freezing in the trained fearful context only on short timescales of a few days after the fearful event, but not in the long term. This is consistent with the idea that context memories may be supported by extra-hippocampal (cortical) brain regions at later times. The acquisition of fear memories also intimately involves the basolateral complex of the amygdala, specifically the lateral and basolateral nuclei.
Researchers found that increasing CREB levels in the basolateral amygdala of rats produced Long term fear memories under a training regimen that typically just produces short-term memories, namely, under massed training regimen. Spaced fear conditioning, where training trials are spaced with rest intervals, is more effective at creating Long term memories than massed fear conditioning, where trials are not intervened with rest intervals.[11]
Paired Associate Learning
Paired-associate learning is a memory paradigm common in humans and other primates that assesses Long term memories formed through object association. In this task, a series of words or pictures are paired and presented to the subject. The subject is then asked to retrieve the other associated pair member at a later time or recognize if a presented pair was in fact truly paired during encoding. Subjects with lesions of the medial temporal lobe show significant impairment in this test.[12]
Recall tests
In this task, human subjects are presented with a list (usually made of words) that they must then recall in any order they wish, typically after multiple learning trials. To evaluate Long term memory, the test is done after a delay period of variable length, which can range from an hour to weeks. Performance is typically measured in terms of the total recalled items on the list. In one study of individuals with uni- or bilateral damage to the frontal lobe, free recall after a one day delay period was significantly impaired in the bilaterally-damaged group compared to the unilaterally-damaged group. This difference was explained in terms of deficits in Long term memory, as well as diminished frontal mechanisms such as attention, motivation, and concentration.[13]
A variant of the free recall task involves a recognition component. In these tasks, subjects are asked to look at pictures of familiar objects, like a banana or pocket watch, and then several days later are shown some of the same images, as well as some novel and lookalike photos. To assess Long term memory, they are asked if they had seen the image before and scored in terms of the total correct responses.
Diseases and conditions that affect Long term memory
Huntington’s disease
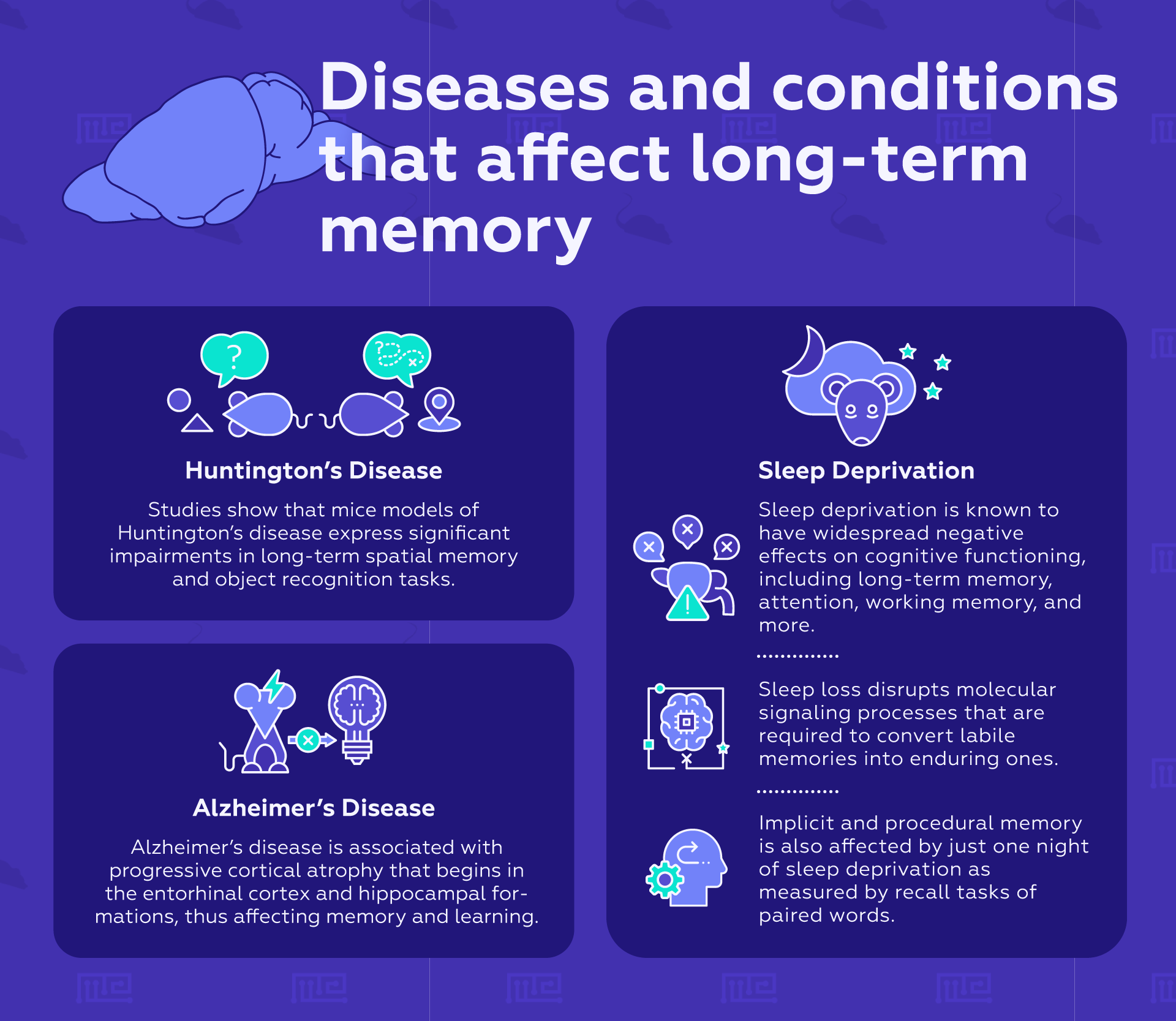
Huntington’s disease is a progressive neurodegenerative disorder caused by a mutation in the coding region of the huntingtin (htt) gene. Although its symptoms are principally motor-related, early cognitive deficits typically appear before the onset of motor symptoms.[14] In HdhQ7/Q111 mutant mice, a mouse model of Huntington’s disease that expresses the htt mutation, diminished hippocampal expression of CREB-binding protein and histone H3 acetylation were causally related to cognitive deficits, implying deficits in memory consolidation. Specifically, these mice show significant impairments in Long term spatial memory and object recognition tasks but spared short-term memory.[15]
Alzheimer’s disease
Alzheimer’s disease is one of the most common neurodegenerative disorders in individuals over the age of 65. One of the hallmarks of Alzheimer’s disease is the formation of misfolded proteins both inside and outside neurons. These neurotoxic proteins include misfolded amyloid-beta, which clump together into amyloid plaques, as well as tau protein, which accumulates into neurofibrillary tangles. The progressive cortical atrophy associated with the disease begins in the entorhinal cortex and hippocampal formations, regions of the brain implicated in memory and learning.
Rodent models of Alzheimer’s disease have mostly focused on how the pathophysiology of the disease affects spatial learning and memory. Researchers commonly utilize the Morris water maze to evaluate Long term spatial memory performance in these animals. In a 2016 study that used a PDAPP genetic model of Alzheimer’s disease, mice were first trained to escape the water to the platform, and then spatial memory retrieval was tested under long delay periods. Memory retrieval was assessed after a seven-day consolidation period alongside concurrent analysis of glucose uptake in the brain. The mice were found to have normal learning and glucose metabolism during the training period, but substantially impaired glucose metabolism and memory retrieval on a test after a seven-day interval. This study suggests that Long term memory tests with long delay periods may be a more sensitive biomarker to early Alzheimer’s disease than short-term memory tests.[16]
Sleep deprivation
Jenkins and Dallenbach were the first to discover that sleep facilitates Long term memory formation in the mid-1920s. In their research, human subjects displayed enhanced memory of verbal syllables when they slept during the period between learning and recall.[17] Numerous other studies have confirmed their results. Sleep deprivation is known to have widespread negative effects on cognitive functioning, including Long term memory, attention, working memory, and more. Sleep, especially slow-wave NREM sleep, plays an important role in memory consolidation. Sleep loss disrupts molecular signaling processes that are required to convert labile memories into enduring ones. Implicit and procedural memory is also affected by just one night of sleep deprivation as measured by recall tasks of paired words.[18]
Drugs that affect Long term memory
Some of the most well-studied drugs that affect Long term memories act on GABA and histamine signaling within the central nervous system. Here we will discuss several drugs that have been shown in humans to disrupt Long term memory processes, including memory consolidation.
GABA-boosting drugs
Alcohol increases GABA activity at the GABAA receptor by acting as a positive allosteric modulator, binding to a site on the receptor and increasing its activity when GABA binds. Alcohol is known to negatively affect encoding, storage, and retrieval of Long term memories.[19] Specifically, alcohol’s effects on GABA and NMDA neurotransmission impair the ability to form new memories by disrupting Long term potentiation and other mechanisms of synaptic plasticity in the hippocampus. In a study of 160 moderate to heavy drinkers, performance on recall and recognition tests of Long term memory were significantly impaired from alcohol intoxication.[20]
Benzodiazepines are a class of drugs commonly used to treat anxiety and sleep disorders. Benzodiazepines also work by increasing GABA signaling at the GABAA receptor. Frequent, high and frequent doses of benzodiazepines are associated with anterograde amnesic properties and Long term memory impairments. This includes disruptions in both memory acquisition and the consolidation of information in Long term memory.[21] A 2004 meta-analysis in humans concluded that benzodiazepines produce a widespread effect on several variables of cognition, including deficits in verbal memory, nonverbal memory, attention, and sensory processing.[22]
Histamine-boosting drugs
Histamine is a neurotransmitter produced primarily in the tuberomammillary nucleus that plays an important role in learning, memory, wakefulness, and circadian processes. Antihistamines are commonly used drugs known to produce drowsiness, but also impair learning and memory and may even increase the risk of dementia in older people.
According to research recently published in Biological Psychiatry, histamine receptor inverse agonists such as thioperamide and betahistine have been shown to promote the retrieval of forgotten Long term object memory without impacting short-term memory. Mice treated with these drugs just once were induced to recall forgotten memories as far out as one week and one month after training. The researchers suggest that the disinhibition of histamine release in the perirhinal cortex facilitates spontaneous activity within these neurons, which in turn promotes memory retrieval. In healthy adults, betahistine treatment similarly promoted enhanced object recognition memory one week after training, specifically for difficult items in the image recognition task.[23]
Conclusion
A properly functioning Long term memory is a foundational part of existence for rodents and humans. Long term memory is impacted by aging, drugs, and a range of common diseases and conditions.
Experimental protocols in humans and rodents have been developed to assess Long term memory performance, including water land-based mazes as well as fear conditioning in rodents, and a variety of recall and recognition tasks in humans.
There is still much to learn about the neurobiological mechanisms behind Long term memory formation, but recent studies utilizing modern techniques have made significant progress in understanding the molecular, systems-level, and behavioral correlates of this memory system.
References
- Lum, J. A., & Conti-Ramsden, G. (2013). Long term memory: A review and meta-analysis of studies of declarative and procedural memory in specific language impairment. Topics in language disorders, 33(4), 282–297. doi:10.1097/01.tld.0000437939.01237.6a
- Kandel E. R. (2012). The molecular biology of memory: cAMP, PKA, CRE, CREB-1, CREB-2, and CPEB. Molecular brain, 5, 14. doi:10.1186/1756-6606-5-14
- Ford, L., Ling, E., Kandel, E.R., and Fioriti, L. (2019). CPEB3 inhibits translation of mRNA targets by localizing them to P bodies. Proc. Natl. Acad. Sci. U.S.A. In review.
- J.S. Stephan, L. Fioriti, N. Lamba, L. Colnaghi, K. Karl, I.L. Derkatch, E.R. Kandel The CPEB3 Protein Is a Functional Prion that Interacts with the Actin Cytoskeleton Cell Rep., 11 (2015), pp. 1772-1785
- Manelis, A., Hanson, C., & Hanson, S. J. (2011). Implicit memory for object locations depends on reactivation of encoding-related brain regions. Human brain mapping, 32(1), 32–50.
- Squire L. R. (2009). The legacy of patient H.M. for neuroscience. Neuron, 61(1), 6–9.
- Tonegawa S., Morrissey M. D., Kitamura T. (2018). The role of engram cells in the systems consolidation of memory. Nat. Rev. Neurosci. 19, 485–498. 10.1038/s41583-018-0031-2
- T. Kitamura, S.K. Ogawa, D.S. Roy, T. Okuyama, M.D. Morrissey, L.M. Smith, R.L. Redondo, S. Tonegawa. Engrams and circuits crucial for systems consolidation of a memory. Science, 356 (2017), pp. 73-78.
- Vorhees, C. V., & Williams, M. T. (2006). Morris water maze: procedures for assessing spatial and related forms of learning and memory. Nature protocols, 1(2), 848–858.
- Attar A, Liu T, Chan WT, Hayes J, Nejad M, Lei K, et al. A shortened Barnes maze protocol reveals memory deficits at 4-months of age in the triple-transgenic mouse model of Alzheimer’s disease. PloS one.
- S.A. Josselyn, C. Shi, W.A. Carlezon Jr., R.L. Neve, E.J. Nestler, M. Davis Long term memory is facilitated by cAMP response element-binding protein overexpression in the amygdala. J. Neurosci., 21 (2001), pp. 2404-2412.
- Sakai K., Miyashita Y. Neural organization for the Long term memory of paired associates. Nature. 1991; 354: 152-155.
- Jetter, W., Poser, U., Freeman, R. B., Jr., &Markowitsch, H. J. (1986). A verbal long term memory deficit in frontal lobe damaged patients.Cortex,22, 229–242.
- Cleret de Langavant, L., Fénelon, G., Benisty, S., Boissé, M. F., Jacquemot, C., & Bachoud-Lévi, A. C. (2013). Awareness of memory deficits in early stage Huntington’s disease. PloS one, 8(4), e61676.
- Giralt A, Puigdellívol M, Carretón O, Paoletti P, Valero J, Parra-Damas A, Saura CA, Alberch J, Ginés S. Long term memory deficits in Huntington’s disease are associated with reduced CBP histone acetylase activity. Hum Mol Genet. 2012 Mar 15;21(6):1203–16.
- Beglopoulos V, Tulloch J, Roe AD, Daumas S, Ferrington L, Watson R, et al. Early detection of cryptic memory and glucose uptake deficits in pre-pathological APP mice. Nat Commun. 2016;7:11761.
- Jenkins JG, Dallenbach KM. Obliviscence During Sleep and Waking. Am J Psychol 605–612, 1924
- Forest G, Godbout R. Effects of sleep deprivation on performance and EEG spectral analysis in young adults. Brain Cogn 2000;43:195–200.
- Baddeley, A., Eysenck, M.W. and Anderson, M.C. (2009). Memory. New York, NY: Psychology Press.
- Nelson, T. O., MeSpadden, M., Fromme, K., & Marlatt, G. A. (1986). Effects of alcohol intoxication on metamemory and on retrieval from Long term memory. Journal of Experimental Psychology: General, 12, 279-287.
- Roth, T., Roehrs, T., Wittig, R., & Zorick, F. (1984). Benzodiazepines and memory. British journal of clinical pharmacology, 18 Suppl 1(Suppl 1), 45S–49S.
- Barker MJ, Greenwood KM, Jackson M, Crowe SF: Cognitive effects of Long term benzodiazepine use: a meta-analysis. CNS Drugs. 2004, 18: 37-48.
- Nomura, H.; Mizuta, H.; Norimoto, H.; Masuda, F.; Miura, Y.; Kubo, A.; Kojima, H.; Ashizuka, A.; Matsukawa, N.; Baraki, Z.; et al. Central Histamine Boosts Perirhinal Cortex Activity and Restores Forgotten Object Memories. Biol. Psychiatry 2018.