
The use of animal models in scientific research has a long history. Many species have contributed to research over the years, and if you asked a group of random scientists to name their animal models, the answers would likely feature a handful of classic examples. Rats, mice, guinea pigs, non-human primates, zebrafish, and even worms and fruit flies would likely dominate that list. However, there are a whole lot more than six species out there with something to offer the research community, and while it may be hard to characterize a new model for widespread use, it can be well worth the effort in the long run. One such animal that is gaining recognition for its potential, particularly in the field of neurodegenerative research, is the sheep.
The Value of Sheep as an Animal Model
Most people think of sheep as a resource for food or wool – not so much scientific models. In many ways, they do not fit the mold of a classical animal model. For example, common animal models such as mice and rats reproduce quickly and have relatively short lifespans. This is helpful from both a scientific and economical view, as the rapid generation time can produce large numbers of subjects for experiments without creating a need for much long-term housing. Sheep, on the other hand, can live for over a decade and have a gestation period of almost five months.[1] These qualities are not generally desirable for an experiment that needs to produce lots of subjects; in the time it would take for one sheep to produce offspring, a mouse could reasonably have more than five different litters. However, the lengthier lifespan of the sheep is exactly why it has become an attractive candidate for specific fields of research, such as neurodegeneration.
Neurodegenerative diseases, like Alzheimer’s Disease, Parkinson’s Disease, and Huntington’s Disease, tend to arise later in life – even severe cases generally do not set in until a person has lived at least a few decades. As a result, the short lifespan of typical lab animals is considered a limitation to neurodegenerative research that relies on those models. When an animal only lives for a couple of years, it is difficult to study the chronic effects of a pathology or treatment. In this way, the approximately ten-year lifespan of a large animal model like the sheep is an advantage because it allows ample time to allow a disease to develop or a treatment to produce measurable effects. Importantly, this increased lifespan does not necessarily create exorbitant costs to maintain a flock of sheep, as they can remain in their natural outdoor environment even while being studied and do not require specialized housing facilities.[2]
The sheep have become especially noteworthy for its potential value as a model for Huntington’s Disease (HD). The genetics of HD is relatively simple: it is an autosomal dominant disorder caused by a trinucleotide CAG repeat in the huntingtin (HTT) gene, and the fact that it is a single-gene disorder renders it at least a little more straightforward in terms of studying and modeling.[2] A transgenic sheep model of HD has been achieved via microinjection of mutant human HTT.[1] Perhaps one of the greatest advantages of creating such a model in sheep is that their brain structure is more similar to the human brain than other more classic lab animal models like rodents. In addition to being considerably smaller than a human brain, some of the most substantial differences between rodent brains and human brains are sites that are of critical importance to the origins of diseases like HD, including the striatum and cortex.[2] Rodents do not have the same distinct striatal nuclei that humans have, nor do they have gyrencephalic cortices, meaning that the rodent brain is smooth while the human brain has a characteristic “wrinkly” appearance due to its convoluted anatomy.
Sheep brains, however, do possess these properties. This provides an added level of confidence when it comes to the translation of neuroanatomical findings from the model animal to the human condition. For example, immunohistochemical experiments in the transgenic sheep model of HD revealed a loss of DARPP32 reactivity in the distinct striatal subregions of the putamen and globus pallidus, a finding that is consistent with pre-symptomatic loss of dopamine receptors in human HD pathology.[1] This more human-like neuroanatomy is not the only advantage that sheep brains have over the more popular model rodent brains – their larger brain size offers a number of practical advantages. Methodologies including PET, EEG, and MRI are difficult to adapt to smaller animals and present a problem regarding the scaling of results from rodent to human. For example, EEGs in humans record from 16-22 electrodes, while EEGs in mice record from only one or two electrodes.[2] As a result, these techniques are more readily adaptable – both in terms of data analysis and physical performance – for a large farm animal like the sheep.
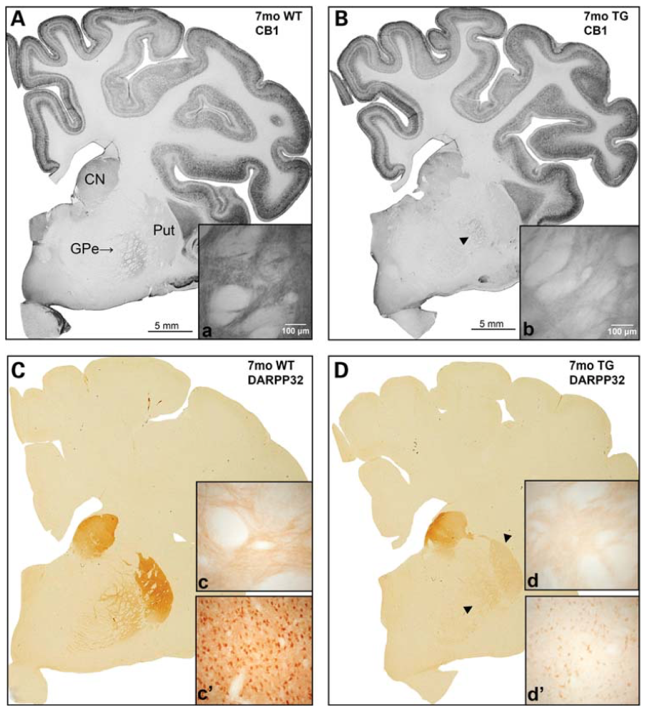
Figure 1: Loss of CB1 and DARPP32 expression in striatum of transgenic HD sheep. [1]
Behavioral Testing in Sheep
In order to fully embrace the sheep as a large animal model for HD, their value must extend beyond neuroanatomy. HD has several hallmark features, among them significant cognitive decline. While sheep are not colloquially known for their intelligence, the recent characterization in behavioral tasks has shown that they possess greater cognitive ability than they have traditionally been given credit for.
Sheep Can Perform Decision-Making Tasks
Given that cognitive function is such an important aspect of HD, a 2011 study[3] sought to characterize the performance of sheep on well-established executive function tasks, specifically reversal learning and attentional set shifting. Both of these tasks are used in the clinic to test the executive function of HD patients. The researchers constructed a set of eight outdoor pens with a series of gates that would guide the sheep through the pens based on its decisions.
In the first set of tasks, the researchers tested the ability of sheep to discriminate between two different colored buckets, one of which contained food and the other did not. Furthermore, an incorrect choice resulted in a brief pause before they could proceed to the next pen and make a new choice.
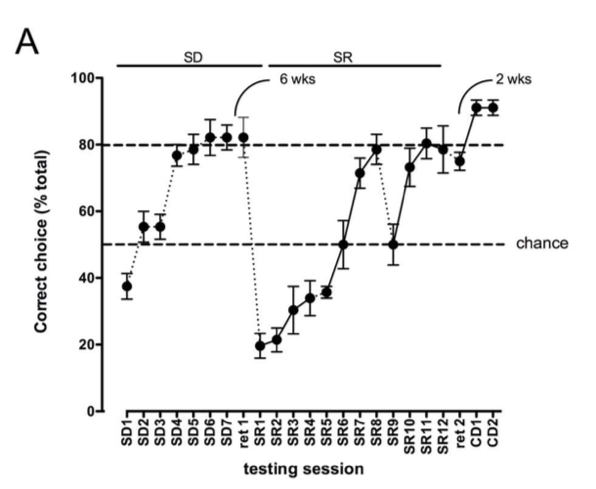
Figure 2: Performance of sheep on a simple discrimination (SD) and simple discrimination reversal (SR) task. [3]
The sheep learned this discrimination easily and could retain that learning for at least six weeks. Eventually, the reward contingency was reversed, and the previously unrewarded bucket became the correct choice. It took some time for the sheep to grasp this reversal; in fact, the sheep seemed reluctant to approach the other bucket, seemingly having learned that an incorrect choice resulted in a quick timeout. They demonstrated a number of emotional reactions to this reversal that resembled anxiety or irritability, including bleating, perseveration around the original rewarded stimulus, and pawing at the researchers. However, they did come around – albeit slowly – and check out the other bucket, and once they found the reward, they ceased their reluctant behavior and were readily making the correct choice by the third day of reversal testing.
In the second set of tasks, the researchers tested the sheep in an attentional set-shifting paradigm in which the sheep must distinguish
between two sets of stimuli – the color and the shape of the bucket – in order to identify where they would receive a reward. They were initially trained to learn that color signaled reward, and eventually that contingency would switch to the previously irrelevant dimension of container shape. Like the discrimination and reversal learning tasks, the sheep were able to discriminate between correct and incorrect choices in the first sets of trials. When the contingencies switch to a new dimension or even within the same dimension but to the previously unrewarded stimulus, the sheep performed significantly worse until they slowly learned the new correct choice. Thus, sheep can be tested successfully in executive function tasks. Furthermore, the sheep seem eager to do so; the researchers noted that before every trial, the sheep demonstrated a number of positive behaviors, including nuzzling the researchers.
Sheep, Like Humans, Are Really Good at Facial Recognition
Humans have evolved to be especially good at recognizing and remembering other human faces. This ability relies on a population of neurons that reside in the temporal and medial prefrontal cortices that are particularly responsive to faces, and a subset of these are more responsive still, to familiar faces. In a task similar to the discrimination task detailed previously, researchers found that sheep were likewise able to distinguish pictures of individual sheep faces that were associated with a food reward.[4] Remarkably, the memory for this association was very strong and persisted for up to 800 days after testing began, thereby implying the existence of a strong network for the storage of these memories.
Leveraging the fact that sheep have a more similar brain size and regional neuroanatomy to humans than other model animals, the researchers recorded from cells in the sheep brain that are analogous to the facial recognition cells in humans. The sheep were presented with two faces, one human and one sheep. The sheep were also divided into three groups: one group was currently in a social group with these individuals, one group was familiar with the individuals but no longer actively interacting with them, and one group had never been familiarized with the individuals. The recordings identified a population of cells that preferentially responded to faces rather than non-face visual stimuli, just as exists in humans. Furthermore, a subgroup of these cells responded preferentially to familiar faces, regardless of whether they were currently interacting with the familiar individual. The sheep even behaved in ways that indicated familiarity, by vocalizing for example. This same cell type did not exhibit a preferential response if the faces were unfamiliar. The study is an
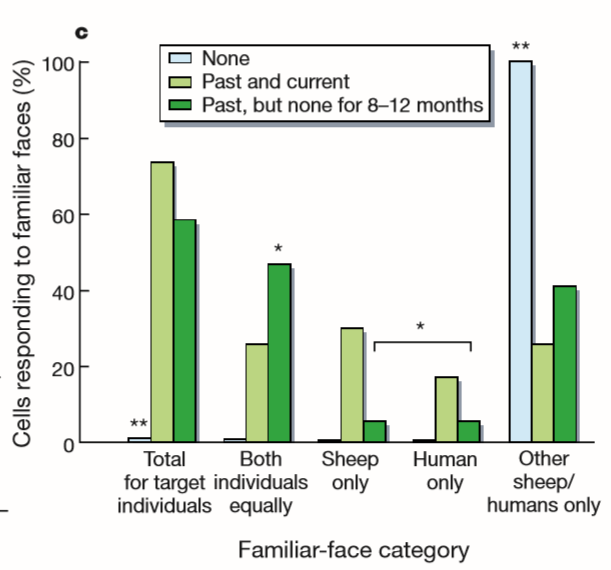
Figure 3: Recordings of cells in sheep brains that respond preferentially to faces.[4]
interesting example of the relevance of neuroscientific data gathered from sheep to the human condition, as it implies the conservation of a defined neural network between the two species.
Measuring Emotions in Sheep
As with any animal model, it is important to rigorously characterize its baseline behaviors before one can fully embrace the model in an experimental setting. One of the greatest challenges of such characterization is determining the emotional or affective state of a given animal. It is hard to resist the temptation to attribute human emotions to animals, and intuition alone will not stand up to scrutiny. However challenging to interpret, it is important to have some grasp on an animal’s affective state if their performance on a given assay is to be interpreted properly. A highly stressed animal, for example, may not perform well on a cognitive test for reasons unrelated to that animal’s relative cognitive ability.
A 2009 study sought to describe the behavioral and physiological characteristics of sheep in three scenarios presumed to evoke different emotional states: separation from the flock was meant to evoke negative emotions, voluntary grooming by a familiar person was meant to evoke positive emotions, and presence in the feeding area was intended to represent a neutral condition. The researchers noted the following behaviors and physiological measurements in each condition:
- Ear Posture: Large farm animals are known to have especially communicative ears. The sheep switched their ear posture most frequently when in the negative condition. The fewest ear posture switches were observed in the neutral condition, with slightly more changes in the positive condition. The sheep had their ears in a forward position the most when in the negative condition, and this proportion decreased as the valence of the situation progressed from negative to neutral and then positive. The opposite trend was seen for ears in the axial position, with the highest proportion in the positive condition and the lowest in the negative condition.
- Eye Aperture: The wideness of the eye changes with arousal state in many species. The sheep’s eyes were open widest during separation from their flock. The aperture decreased in the neutral condition and was the lowest in the positive condition.
- Heart rate Variability: Variation in the sheep’s heart rate was measured by inter-heartbeat interval (R-R interval) and root mean square of successive inter-heartbeat interval difference (RMSSD). Both of these measures increased as the situational conditions became more positive, the lowest when separated and the highest when being groomed by a familiar handler.
- Perspiration: Sensors measured body surface humidity and temperature of the sheep during testing. Neither of these measures appeared to be influenced by the emotional valence of the situation.
- Respiration: The rate of respiration was measured with a belt around the abdomen that would move during inhalation and exhalation cycles. Although there was a trend towards decreased respiration rate as the conditions became more positive in emotional valence, the result was non-significant
As previously mentioned, affective state can impact performance on cognitive assays. This is why, for example, it is common practice to structure a series of behavioral assays, to begin with, the least stressful assay and end with the most stressful assay, as the stress induced by one behavioral test can impact the results of subsequent tests. It is therefore important to understand the influence of a sheep’s affective state on decision making or other cognitive domains in order to accurately interpret their performance in a given context.
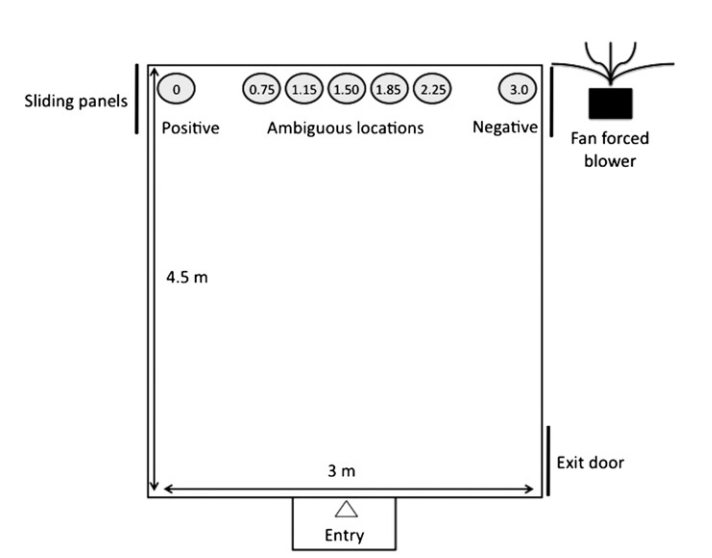
Figure 4: Schematic of the testing arena for assessing the judgement biases of sheep.[6]
One group of researchers dissected this problem by investigating the impact of long-term stressors on the judgment biases of sheep.[5] Young lambs were trained to approach a bucket in a specific corner of an arena with a food reward. The same sheep were also trained to associate a bucket in the opposite corner with an aversive stimulus – a fan-forced blower that would turn on when approached. When the sheep had learned this association, half of the sheep were subjected to a long-term regimen of aversive, random events that are common to life on a farm, like dogs, ear tattooing, shearing, restraint, or being transported on a truck.
The sheep were then placed into the arena and presented with a bucket in an ambiguous location – not quite the food corner, but not
quite the corner with the fan, either. Interestingly, sheep in either group tended to approach buckets in ambiguous locations at the same rate, save for one specific ambiguous location (1.15m from the positive bucket) that the stressed sheep approached less often. This could indicate some degree of negative judgment bias resulting from the stressors, as their decline in approaches to that ambiguous location could be interpreted as a pessimistic response, but it is not conclusive. Stressed sheep also did not exhibit significant differences from control sheep on a series of emotional reactivity assays. Thus, it is possible that exposure to chronic stress has an impact on the judgment biases of sheep, but the effects of stress on the cognition and emotional reactivity of sheep requires more investigation before the results can be faithfully interpreted.
All things considered, the benefits of adopting more large animal models in neuroscience research may well outweigh the costs – of both time and money – involved in characterizing a new model animal. That the data obtained from sheep may be more easily interpreted and applied to human studies, particularly in the field of neurodegeneration, adds considerable value to the concept of utilizing sheep models on a greater scale. However, there is still a lot of work to be done in terms of characterizing the baseline behaviors of sheep, as well as their reactions to common experimental manipulations like stressors, before they can be adopted with full confidence as a behavioral model.
References
- Jacobsen, J.C., Bawden, C.S., Rudiger, S.R., McLaughlan C.J., Reid, S.J., Waldvogel, H.J., MacDonald, M.E., Gusella, J.F., Walker, S.K., Kelly, J.M., Webb, G.C., Faull, R.L.M., Rees, M.I., and Snell, R.G. (2010). An ovine transgenic Huntington’s disease model. Human Molecular Genetics 19(10): 1873-1882.
- Morton, A.J. and Howland, D.S. (2013). Large genetic animal models of Huntington’s disease. Journal of Huntington’s Disease 2: 3-19.
- Morton, A.J. and Avanzo, L. (2011). Executive decision-making in the domestic sheep. PLoS One 6(1): e15752.
- Kendrick, K.M., da Costa, A.P., Leigh, A.E., Hinton, M.R., and Peirce, J.W. (2001). Sheep don’t forget a face. Nature 414: 165-166.
- Doyle, R.E., Lee, C., Deiss, V., Fisher, A.D., Hinch, G.N., and Boissy, A. (2011). Measuring judgement bias and emotional reactivity in sheep following long-term exposure to unpredictable and aversive events. Physiology & Behavior 102: 503-510.
- Reefmann, N., Wechsler, B., and Gygax, L. (2009). Behavioural and physiological assessment of positive and negative emotion in sheep. Animal Behaviour 78: 651-659.