





The Win Stay maze is based on in-shift task as done by Berke et al with rats, whereby the dorsal hippocampus and striatal cells are measured during the task with overwhelming activation of the hippocampal place cells. The dorsal/lateral striatum are more important for learning and choosing actions in body-centered (egocentric) coordinates, such as a left-turn response (Brasted et al. 1997; Cook and Kesner 1988), whereas the hippocampus is more important for remembering particular spatial locations defined by arrays of external cues (allocentric coordinates;Morris et al. 1982).
When a behavioral task forces an animal to make use of one form of information over the other, inactivation of striatum increases use of a hippocampal-dependent “place” strategy, whereas inactivation of hippocampus increases use of a striatum-dependent egocentric “response” strategy. Thus, they are in contrast to each other.
This maze utilizes this anatomy specifically for the win shift task.
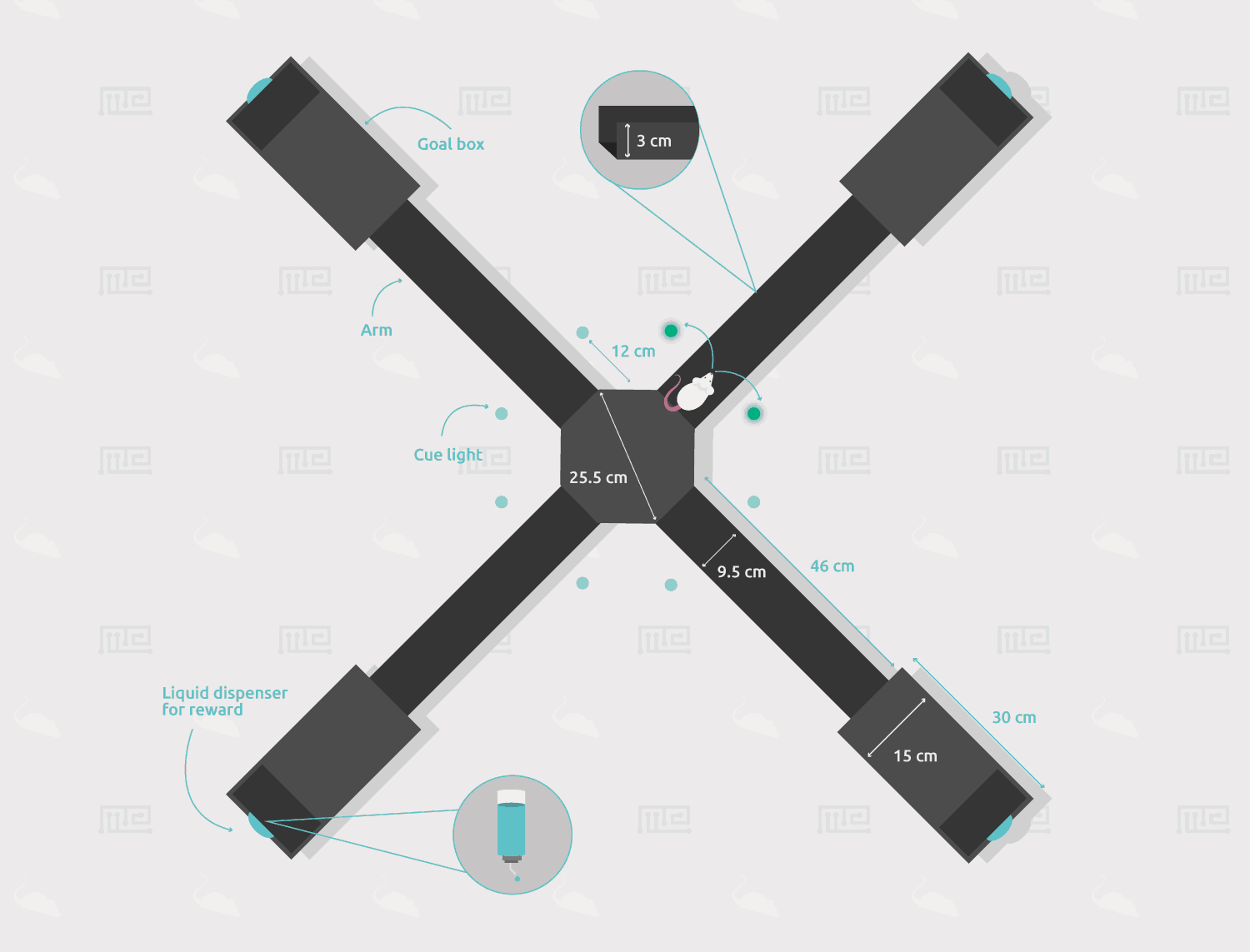
-Maze: Acrylic plus maze, matte black, elevated 71 cm from the floor. Central octagon (25.5-cm diameter), four arms (46 × 9.5 cm), and four goal boxes at the end of the arms (30 × 15 cm). 3cm guard rails.
-Cue lights (NOT) included in the order: Green LEDs on stalks, located 12 cm into each arm) indicated the rewarded arm on each trial
-Liquid dispensers (4) are included in the order.
Price & Dimensions
Mouse
$ 4990
Per Month- Central Platform
- 4 Arms
- 4 goal boxes
- 3cm high railings
- 4 lickometers, controlled with Conductor Software



The Y maze with a social interaction cage was first described by Weber-Stadlbauer et al and replicated by Vuillermont in analyzing social interaction choices in rodents. Sizes are standard for mice and rats, but with the key modification being pair of rectangular wire grid cages. In the protocol, one wire cage contained an unfamiliar rodent (usually same sex as test mouse), and the other one contained a “dummy object.” The Mazeengineers modification of the Y maze includes two removable rectangular wire grid cages for social interaction. This modification is an add on to the regular Y maze and requires separate purchase of the Y maze.
Documentation
Introduction
The Win-Stay maze is used in the investigation of neural activity in the hippocampus and striatum. The dorsal/lateral striatum plays an important role in learning and choosing actions based on egocentric cues while the hippocampus enables learning spatial locations based on a range of external cues. Abnormalities in the neuronal pathways are believed to be an underlying factor in many neurological and psychiatric disorders that include Parkinson’s disease, obsessive-compulsive disorder, and drug addiction.
The Win-Stay maze rewards the subject for choosing an arm based on the cues rather than their previous choices. The Win-Stay maze enables comparison of win-stay and win-shift strategies. In subjects with damage to the sensorimotor areas of the striatum, a decreased ability in performing win-stay tasks can be observed, while hippocampal damage results in decreased ability to perform the win-shift task. Further, it has been observed that inactivation of striatum increases the use of a hippocampal-dependent place strategy in tasks that force the animal to use one form of information over another. On the other hand, inactivation of the hippocampus increases the use of a striatum-dependent egocentric response strategy
The Win-Stay maze consists of an Elevated Plus Maze that has each arm-end equipped with a goal box and LED lights that serve as stimulus cues. Subjects are often implanted with electrodes to observe and record place cell firings as they perform on the Win-Stay maze. Other mazes used in the observation of neural activity include the Hairpin Maze and the Continuous Angled T-maze. The Radial Arm Maze has also been used to investigate win-stay and win-shift strategies (Berke et al., 2004).
Apparatus & Equipment
The Win-Stay Maze utilizes a matte finish, opaque elevated plus maze. The central arena is an octagon of diameter 25.5 cm that has four arms of length 46 cm and width 9.5 extending perpendicularly from it. The ends of each arm are equipped with goal boxes of dimensions 30 x 15 cm. The arms and arena have a 3 cm high lip along their edges to prevent the subject from falling over. At 12 cm into each arm, LED lights on either side are placed to serve as visual cues. The maze is elevated to a height of 71 cm from the floor.
Training Protocol
The apparatus is cleaned before and after every trial to limit the influence of any lingering stimuli. Video and tracking device, such as the Noldus Ethovision XT, is used for tracking and data collection. Ensure the apparatus is appropriately lit during the trials. The apparatus is surrounded with thick black curtains and visual cues, if any, are positioned.
Prior to the experiment, the subjects are implanted with tetrodes (12.5-μm wire) into both the dorsal hippocampal CA1 pyramidal layer and the striatum to observe and record neural activity. The subjects are given at least a week to recover from surgery before beginning behavioral testing. In preparation for the Win-Stay maze, restrict water consumption of the subjects prior to the training.
Win-Stay Maze Task
Place the subject in one of the arms to initiate the task. Once the subject reaches the central arena
Light up the LEDs of the goal arm. The selection of the goal arm should be pseudorandom though the selection must satisfy two conditions: the arm that the subject is currently on is never the selected arm, and if the subject has made the same egocentric choice on the previous two trials, then the subject is not cued to make the same movement a third time in a row. Once the subject reaches the right goal, dispense the water via water dispensers. The trials are conducted in a continuous manner without any intervals between each trial. Thus, for each trial, the previous goal arm becomes the start arm.
Investigation of striatal versus hippocampal representations during Win-Stay Maze task
Adult, male Long-Evan rats were trained on the continuous win-stay maze task motivated by water restriction. Subjects were implanted with tetrodes into both the dorsal hippocampal CA1 pyramidal layer and the striatum, or a bundle of stereotodes was implanted in multiple subregions of the striatal. Rats performed continuous win-stay task using visual discrimination over 270 sessions during which conflict trials (when the subject was randomly cued to return to the arm they had visited on the immediately preceding trial) were also performed. It was observed that the rat performances progressively improved over trials. Though initially the subject utilized spatial foraging strategy held dominance during the trials, subjects were observed to progressively switch to win-stay strategy almost exclusively based on the analysis of conflict trials. Neural activity observation revealed an overwhelming activation of hippocampal neurons at single spatial locations while striatal projection neurons showed activity at discrete points significantly during choices or following reward. However, place-cell-type firing could not be observed. (Berke et al., 2009)
Data Analysis
The following parameters can be recorded during the Win-Stay maze task.
- Latency to initiate the task
- Latency to enter an arm
- Latency to reach the goal arm
- Number of correct choices
- Number of errors
- Path taken
- Velocity
Data analysis of neural activity can include the following.
- Correlation procedures
- Directional tuning
- Egocentric scores
- Population vector construction
- Rate Maps
- Spatial autocorrelation analysis
- Spatial firing patterns
- Trajectory replay
- Turning point analysis
Strengths & Limitations
Strengths
The Win-Stay maze can be easily adapted from an Elevated Plus Maze to investigate the competitive and corporative interactions between the hippocampus and striatum. The four-arm maze reduces the complexity associated with data collection and observation in comparison to the Radial Arm Maze. The conflict trials in the Win-Stay maze enable evaluation of conflict between win-stay and win-shift strategies early in learning. Further, the symmetrical design enables assessment of allocentric encoding by observation of neuron firing on the four arms. The Win-Stay maze can be effectively used in the assessment of the effects of lesions and drug treatments on learning and memory of the subjects. The task can also be used in evaluation learning and memory abilities and in the investigation of the underlying causes in neuropsychiatric and neurodegenerative disorders.
Limitations
Repeated trials on the Win-Stay maze can potentially lead to experimental fatigue. The elevated nature of the maze can cause anxiety and stress in the subjects. Also, the open space may also contribute to these emotions, thus having a potential influence on the task performance. The subject’s handling and transportation to the maze could also potentially affect the data and its behavior in the maze. Presence of any unintended stimuli or cues, could also potentially influence the subject’s performance. The Win-Stay maze task can be time-consuming. The natural biases of the subjects against or towards a certain arm cannot be eliminated despite the symmetrical arrangement of the arms. The flashing light cue is a strongly directing behavior and might, therefore, evoke correspondingly large changes in neural activity.
Summary & Key Points
- Win-Stay maze task is used to evaluate neural activity associated with memory and learning.
- The maze is used to train the subject to perform continuous win-stay task using visual discrimination.
- The conflict trials allow investigation of win-stay and win-shift strategies.
- In comparison to the Radial Arm Maze, the Win-Stay maze reduces the complexity of data collection and observation.
- The flashing light cue strongly directs the behavior of the subjects in the task.
- Repeated trials can lead to experimental fatigue.
- The elevation and open nature of the maze can cause anxiety and stress in the subject. Subject’s own bias can also influence performance.
References
Berke, J. D., Breck, J. T., & Eichenbaum, H. (2009). Striatal Versus Hippocampal Representations During Win-Stay Maze Performance. Journal of Neurophysiology, 101(3), 1575–1587.
Berke, J. D., Okatan, M., Skurski J., Eichenbaum, H. B. (2004). Oscillatory entrainment of striatal neurons in freely moving rats. Neuron 43, 883–896
Blazquez, P. M., Fujii, N., Kojima, J., Graybiel, A. M. (2002). A network representation of response probability in the striatum. Neuron 33, 973-982