












Description
The Hairpin maze is a unique maze has been described in the literature for studying grid place cells. Mouse and rats run through interconnected compartments of equal shape and orientation in 10 equal length alleyways.
Available in Black/Blue/White/Grey/Red/Clear
A removable version is available allowing conversion into an open field box as well as insertion of truncated arms.
Price & Dimensions
Static Model
Mouse
$ 1800
Per Month- 60 cm × 60 cm
- 10 equal length alleyways
- Width: 6cm
- Available in Black/Blue/White/Grey/Red/Clear
Rat
$ 1900
Per Month- 100 cm × 100 cm
- 10 equal length alleyways
- Width: 10cm
- Available in Black/Blue/White/Grey/Red/Clear
Removable Model
Mouse
$ 2200
Per Month- 60 cm × 60 cm
- 10 equal length alleyways
- Width: 6cm
- Available in Black/Blue/White/Grey/Red/Clear
Rat
$ 2300
Per Month- 100 cm × 100 cm
- 10 equal length alleyways
- Width: 10cm
- Available in Black/Blue/White/Grey/Red/Clear
Documentation
Introduction
The Hairpin Maze provides a more realistic environment as opposed to the Open-field. The interconnected zig-zag alleyways enable compartmentalization of the maze that effectively mimics the complex and nested environments of the subject.
The hippocampus plays an important role in learning, spatial memory, and navigation, making it an extensively studied part of the brain system. The discovery of place cells (O’Keefe & Dostrovsky 1971) and grid cells (Hafting et al., 2005) have led to better understanding of how the neural network works as a mapping system. In search of gaining further insights on how the brain maps spaces, the Hairpin Maze serves as a unique apparatus to assess entorhinal-hippocampal representations of places, routes, and associated experiences.
Using rodent model, the hairpin maze helps determine spatial orientation and mapping taking place in different environments based on the firing of the grid cells, the place cells, and the head direction cells. Place cell firing leads to cognitive mapping based on the allocentric place fields. The place fields can undergo “complete remapping” when the subject is moved to a different environment (Muller & Kubie, 1987) or show lesser changes (“rate remapping”) when minor changes to the environment are made (Fyhn et al., 2007), suggesting their sensitivity towards visuospatial information. On the other hand, grid cells have an iterative triangle array of multiple firing fields that are unaffected by spatial and non-spatial cues (Fyhn et al., 2007). The grid cells aid the place cells in path integration to allow tracking of spatial location even in the absence of any cues. The head direction cells are found in many parts of the brain but substantially more intermingled with deeper layers of the grid cells. These cells are dependent on the vestibular system (Blair & Sharp 1996) and fire in relation to the head direction specificity independent of a functional hippocampus (Golob et al., 1990).
Apparatus and Equipment
The Hairpin Maze apparatus consists of a 150 x 150 x 50 cm arena with nine walls of dimensions 135 cm x 30 cm x 1 cm placed at equidistance (14 cm) to create a zig-zag alleyway. The apparatus is constructed using acrylic material and can be opaque or transparent. The apparatus is also available with removable walls for easy conversion into an Open-field arena.
Training Protocol
The apparatus is cleaned before and after every trial to limit the influence of any lingering stimuli. Video and tracking device, such as the Noldus Ethovision XT, is used for tracking and data collection. The apparatus is surrounded by thick black curtains and visual cues, if any, are positioned.
Before to the experiment, tetrodes are inserted above the dorsocaudal part of medial entorhinal cortex (MEC), 4.5 mm lateral to the midline and 0.2–0.3 mm anterior to the transverse sinus, at an angle of ~10° in the anterior direction in the sagittal plane to record neural activity (Derdikman et al., 2009).
Each daily session consists of an initial Open-field run, two Hairpin Maze runs, and a final Open-field run. It is also important to ensure that the subjects are allowed resting period between each run.
Pre-training in the Open-field Arena
The subjects are introduced to the Open-field arena and food crumbs are randomly thrown around the surface for them to collect. During this task, a white cue card (95 x 45 cm) is placed 110 cm above the floor on the curtain. Once the subject regularly covers the entire area of the Open-field in the 20-minute trial, the training on the Hairpin Maze is initiated. If using a grooved Open-field, a removable linoleum mat is placed in the arena.
Hairpin Maze Task
The subject is introduced into the Hairpin Maze from the start position. On alternating trials, the subject is trained to run from east to west and vice versa without interruption. During the trial, food crumbs can be used to motivate the subject to explore the maze. The trial lasts for 20 minutes. After a resting period, the subject is once again placed in the Open-field arena for a 20-minute run.
Short-cut Hairpin Maze Task
Two central arms are replaced with 85 cm length walls and a barrier wall (29 cm) is placed perpendicular to the run direction 100 cm from the outer wall. The training and testing are done as mentioned earlier (Derdikman et al., 2009).
Virtual Hairpin Maze Task
The Virtual Hairpin Maze task involves the Open-field arena. As opposed to scattering food randomly in the arena, the food rewards are delivered at the successive west to east or vice versa positions, one at a time at the north and the south walls in an alternating north-south sequence. The distance between the location of the food rewards mimicked the width of the Hairpin Maze alleys. The resulting north-south run pattern is similar to the Hairpin Maze path (Derdikman et al., 2009).
Sample Data
The analysis is performed based on the cell firing data, position data and path for both Open-field and Hairpin Maze runs. A separate analysis is performed for westbound trajectories (path traversal from arm 9 in the east to arm 2 in the west without returning to arm 9) and eastbound trajectories (path traversal from arm 2 to arm 9 without returning). Outer arms are excluded from the analysis due to long durations of inactivity by the subject. Cell firing is recorded using the recording electrodes placed in the MEC and position estimates are based on the tracking of the LED on the headstage.
Data analysis include the following.
- Rate Maps
- Spatial autocorrelation analysis
- Directional tuning
- Population vector construction
- Correlation procedures
- Turning point analysis
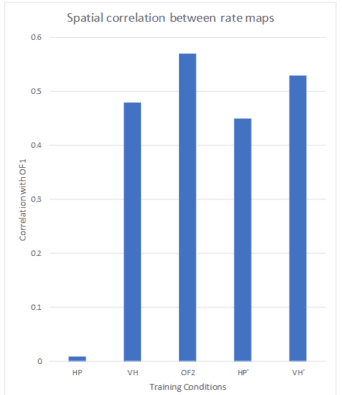
The following sample graph depicts the spatial correlation between rate maps of the first Open-field (OF1), the Hairpin Maze task (HP), the Virtual Hairpin (VH) and the second Open-field (OP2) run.
Strengths & Limitations
Strengths
The Hairpin Maze’s interconnected zig-zag alleyways effectively mimic the complex and nested environments of the subject. Since it closely reflects the natural habitat of the subject, it allows observation of grid cells, place cells and head direction cells firing as they would in the real environments. It is a unique, simple and effective maze for evaluating entorhinal-hippocampal representations of places, routes, and associated experiences.
Limitations
Head direction cells are independent of a functional hippocampus to express their head direction specificity. Their role, if any, in the network mechanisms that induce remapping in the hippocampal-entorhinal complex still requires further understanding. The subject’s handling and transportation to the maze could also potentially affect the data and its behavior in the maze. The Hairpin Maze task can be time-consuming.
Summary and Key Points
- The Hairpin Maze serves as a unique apparatus to assess entorhinal-hippocampal representations of place, routes, and associated experiences.
- The Hairpin Maze’s multiple alleyways present an environment with changing stimuli and directions, closely reflecting the subject’s natural habitat.
- The maze allows observation of grid cells, place cells and head direction cells firing as they would in the real environments.
- The maze serves as an effective way to determine the neural representation of spatial orientation and mapping.
References
Derdikman, D., Whitlock, J. R., Tsao, A., Fyhn, M., Hafting, T., Moser, M., & Moser, E. I. (2009). Fragmentation of grid cell maps in a multicompartment environment. Nature Neuroscience, 12(10), 1325-1332.
Fyhn M, Hafting T, Treves A, Moser MB, Moser EI (2007). Hippocampal remapping and grid realignment in entorhinal cortex. Nature. 446(7132):190-4.
Golob, E. J.; Taube, J. S. (1999). Head direction cells in rats with hippocampal or overlying neocortical lesions: evidence for impaired angular path integration .The Journal of Neuroscience. 19 (16): 7198–7211
Hafting T, Fyhn M, Molden S, Moser MB, Moser EI (2005). Microstructure of a spatial map in the entorhinal cortex. Nature. 436(7052):801-6.
Muller, RU; Kubie, JL (1987). The effects of changes in the environment on the spatial firing of hippocampal complex-spike cells. J Neurosci. 7: 1951–68.
O’Keefe, J., and Dostrovsky, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 34, 171–175
Whitlock, J. R., & Derdikman, D. (2012). Head direction maps remain stable despite grip map fragmentation. Frontiers in Neural Circuits, 6.
Blair, H. T.; Sharp, P. E. (1996). Visual and vestibular influences on head-direction cells in the anterior thalamus of the rat. Behavioral Neuroscience. 110 (4): 643–660. doi:10.1037/0735-7044.110.4.643.