
Nootropics represent a class of drugs or dietary supplements aimed at enhancing cognition. Primarily, nootropics target the cognitive domains of executive function, memory and creativity. Drugs which have been classified as confirmed or potential nootropics are often those initially investigated for the amelioration of cognitive dysfunction in aging and neurodegenerative disorders, as well as the treatment of attention or learning deficits in such conditions as attention deficit hyperactivity disorder (ADHD) or autism spectrum disorders. These include the pharmaceutical amphetamines, eugeroics (such as modafinil), as well as the stimulants caffeine and nicotine. Dietary supplements which have been embraced as potential or confirmed nootropics range from amino acids such as D-aspartic acid, L-theanine and beta-alanine to those of the plant domain such as ginseng and Lion’s Mane mushroom.
Categorizing the Nootropics
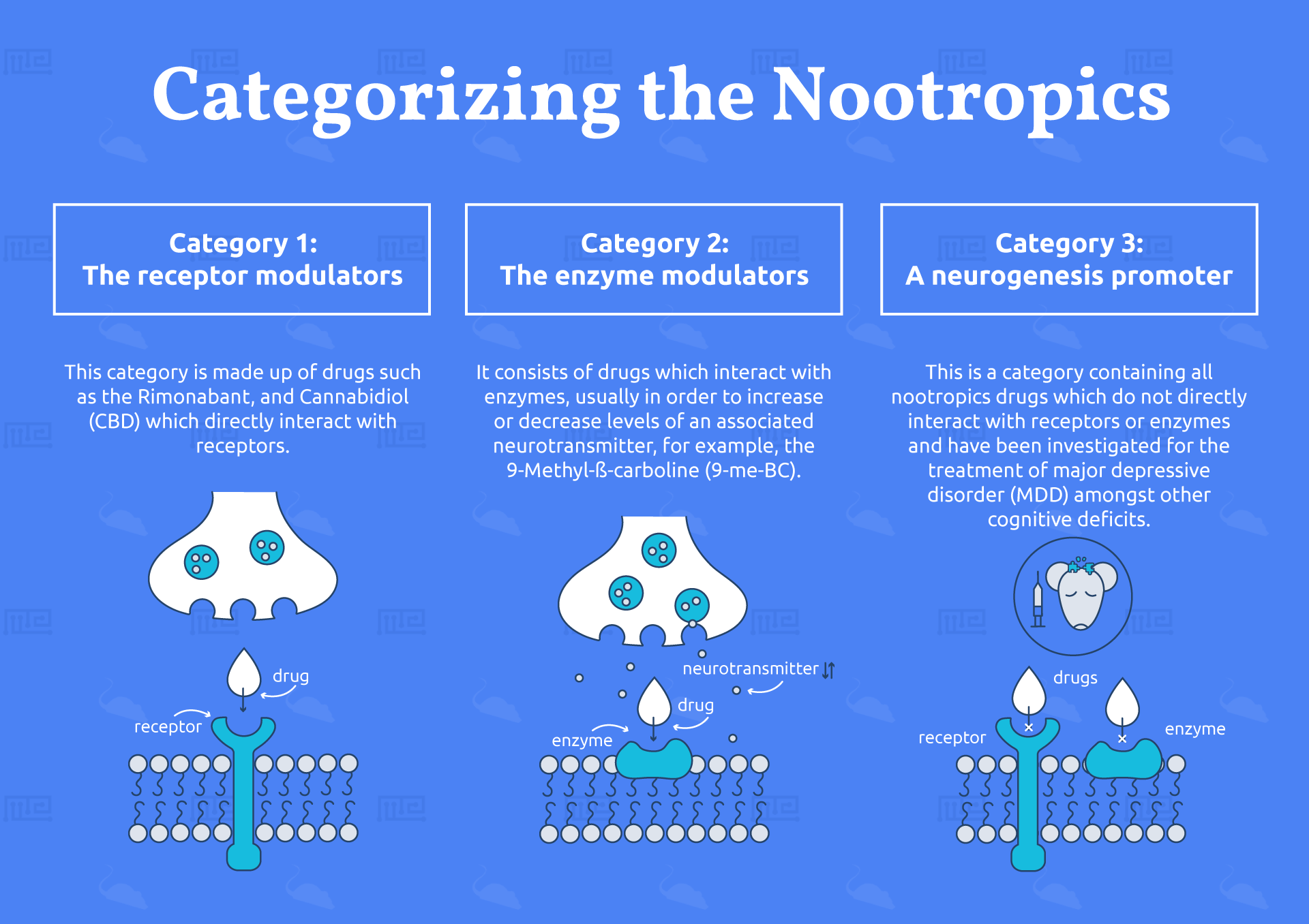
Broadly, nootropics fall into three pharmacological categories as delineated by the Swiss researchers Wolfgang Froestl, Andrea Pfeifer and Andreas Muhs.
Category 1
This category is made up of drugs which directly interact with receptors, including the classic nootropic piracetam (the inventor of which, Corneliu Giurgea, also coined the term nootropic) and the stimulants mentioned above.[1]
Category 2
The second category is comprised of drugs which interact with enzymes such as acetylcholinesterase, the most well-known of which is probably L-Huperzine, a moss-extract long-known for its nootropic properties.[2]
Category 3
The third category contains all of the nootropics which do not directly interact with either receptors or enzymes, but rather function through a variety of other mechanisms, such as preventing ß-amyloid or tau tangles in neurodegeneration, peptide or hormone modulation, or those which display antioxidant properties.[3]
Here, we will discuss several putative nootropics of interest, spanning these categories with a special focus on lesser-known, novel approaches to cognitive enhancement. It is important to note that most putative nootropic drugs have been developed for use in individuals with cognitive deficiencies, rather than healthy people seeking to enhance their mental acuity, memory or executive functions. Very few scientific findings regarding nootropic efficacy exist in humans, and even less in healthy, non-compromised subjects. Thus, the field of nootropics is largely driven by extrapolative findings rather than robust direct evidence, and should be considered in this light.
Category 1: The receptor modulators
Rimonabant
While cannabis and cannabinoids are most commonly associated with cognitive impairment and intoxication, the endocannabinoid system through which they work is an attractive potential target for cognitive enhancement. One drug, though discontinued as a pharmaceutical treatment as a result of potential concerns regarding its propensity to induce depression symptomology in a subset of patients, is of particular interest as a potential nootropic. SR 141716, better known as Rimonabant, is an inverse agonist at the type-1 cannabinoid receptor (CBR1), and thus belongs to the first pharmacological category of nootropics.
First identified as a potential cognitive enhancer in 1996 by Terranova and colleagues at the French pharmaceutical development firm, Sanofi, Rimonabant was found to be of interest due to its ability to reverse the inhibition of hippocampal synaptic plasticity caused by the CBR1 agonist, WIN 55,212-2.[4] This lead the researchers to test the drug alone in the social recognition test, a common measure of social memory.[5] Indeed, Rimonabant was found to enhance social memory in a dose-dependent manner, a finding which would lead these and other researchers down a long path of discovering the cognitive enhancing effects of the drug.
Using the radial arm maze, multiple groups of researchers have found that Rimonabant enhances spatial memory. A study from the year 2000[6] and another, conducted in 2003,[7] examined the drug’s ability to improve spatial memory in two variants of the test, both of which resulted in enhanced performance. Interestingly, the latter of these two studies found that the drug was specifically effective in testing the memory function following a prolonged inter-trial interval, suggesting that it enhances long-term memory consolidation. A similarly acting drug developed by the pharmaceutical company Pfizer, named CE, confirmed this finding by testing rats at an even longer delay than previously investigated with Rimonabant, and also found enhanced spatial memory consolidation.[8]
Rimonabant has also been shown to improve memory in a host of other conditions. Researchers in 2005 tested the effects of the drug on memory consolidation by giving it to mice either before or after the training sessions for memory in an elevated T-maze.[9] Rats treated with Rimonabant showed significantly improved performance when tested either 1 or 7 days after the training session, as compared to control animals. Moreover, memory deficits induced by administration of the drug, scopolamine, were effectively reversed by this Rimonabant treatment. Rescue of other memory deficits by rimonabant include, amnesia induced by injection of beta-amyloid fragments,[10] thought to be one of the peptides involved in the development of alzheimer’s disease and its associated memory dysfunction.
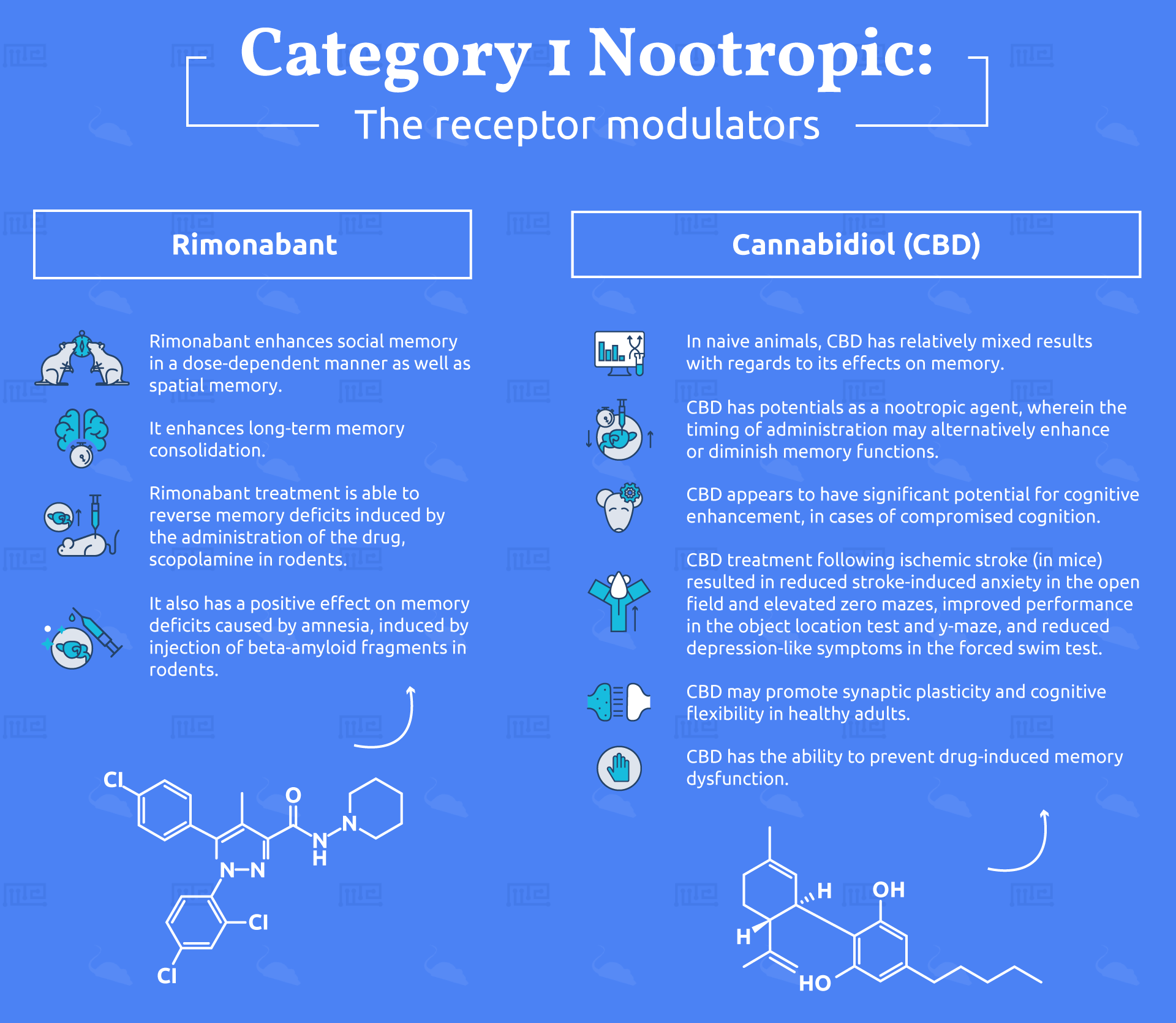
Cannabidiol (CBD)
While Rimonabant was recalled as a pharmaceutical, worldwide in 2008, due to an unexpected increase in depressive symptoms and other side-effects in the larger population, interest in modulation of the cannabinoid system as a potential pharmacological target was only just beginning. One drug of recent interest and popularity in this regard is the phytocannabinoid, CBD. CBD is a naturally occuring cannabinoid, found in the cannabis sativa plant, and has recently been approved by several countries’ drug governing bodies for the treatment of select forms of epilepsy. While the mechanism of action remains unknown,[11] a great deal of attention has come to the drug following its approval by the FDA, along with the current renaissance of cannabis-oriented research.
In naive animals, CBD has relatively mixed results with regards to its effects on memory. In fact, several studies point to CBD’s ability to disrupt memory consolidation when given immediately after a training session in the fear-conditioning task[12-14] or following conditioned-place preference training.[15] However, when given during, rather than after, a training session, CBD appears to enhance memory consolidation in a fear-conditioning paradigm.[16] These findings highlight the specificity of CBD’s potential as a nootropic agent, wherein the timing of administration may alternatively enhance or diminish memory functions.
In contrast to the mixed findings in naive or otherwise, healthy animals, CBD appears to have significant potential for cognitive enhancement, in cases of otherwise, compromised cognition. For instance, following ischemic stroke, mice (and humans) have been shown to exhibit significantly impaired emotional regulation and memory function, accompanied by inflammatory-induced reductions in cortical mass. Researchers in Brazil published, in 2017, their findings on the treatment of these post-stroke conditions with CBD.[17] Using a variety of tasks including the open field, elevated zero maze, y-maze, object location test and forced swim test, they found that CBD treatment following ischemic stroke resulted in reduced stroke-induced anxiety in the open field and elevated zero mazes, improved performance in the object location test and y-maze, and reduced depression-like symptoms in the forced swim test. These findings were accompanied by a reduction in hippocampal neurodegeneration and inflammation, and in fact, an increase in hippocampal neuroplasticity.
In humans, two studies, published in 2013, highlight the potential for CBD as a nootropic drug in two different manners. First, healthy subjects were given 32mg of CBD, in a double-blind, placebo-controlled study, in order to assess its potential effects on fear memory extinction.[18] Individuals who received the drug exhibited more robust levels of extinction following stimulus association and subsequent dissociation than those who received the placebo. These findings indicate that CBD may promote synaptic plasticity and cognitive flexibility in healthy adults. Cognitive flexibility, in particular, is a key aspect of learning, planning and decision making, which makes it of great importance with regards to nootropic efficacy.
Next, a separate research group sought to study the effects of CBD on the cognitive impairment, anxiety and psychotic symptoms associated with administration of cannabis’ primary psychoactive component, ∆9-THC. To do so, they pre-administered either 600mg of oral CBD or a placebo to individuals prior to their ∆9-THC exposure.[19] In this case, CBD-pre-treated subjects exhibited significantly lower levels of anxiety and psychotic-like symptoms than their placebo-dosed counterparts and, more importantly, less memory dysfunction. When tested for episodic memory, subjects given a placebo prior to their THC administration showed a significant impairment while those given CBD performed on par with the control group given no THC, thus highlighting CBD’s ability to prevent drug-induced memory dysfunction.
Category 2: The enzyme modulators
The second category of nootropics is comprised of those which interact with enzymes, usually in order to increase or decrease levels of an associated neurotransmitter. One such compound which has garnered significant attention amongst nootropics researchers is 9-Methyl-ß-carboline (9-me-BC). First identified as a potential neuroprotective agent, in 2008, by researchers in Germany,[20] 9-me-BC was initially found to be of interest due to its ability to upregulate tyrosine hydroxylase, the enzyme responsible for converting the amino acid L-theanine into the dopamine precursor, L-DOPA. This crucial step in the synthesis of dopamine is known to be downregulated or reduced in a variety of neurodegenerative disorders,[21] including Alzheimer’s disease.[22] Thus, after discovering its capability to increase TH levels, 9-me-BC quickly became a subject of interest for the treatment of dopaminergic disorders, many of which are associated with significant memory and learning problems, thus making it equally interesting as a potential nootropic agent.
Following up on their discovery, this same group of researchers tested the effects of 9-me-BC in vivo, by administering the drug intraperitoneally to rats for either 5 or 10 days, and testing their performance in the well-known spatial learning task, the radial arm maze.[23] Additional tests of neuroanatomy and neurotransmitter levels were also performed on these rats, which were compared to both their non-treated and saline-treated counterparts. Importantly, the 9-me-BC-treated rats were otherwise healthy, meaning that the results of the study can be extrapolated to potential nootropic efficacy in healthy humans, rather than, as is often the case, surmising potential effects based off of studies normally performed in either animal models of cognitive disorder, or in humans with reduced mental function. Remarkably, the 9-me-BC treatment significantly improved radial arm maze performance in the rats after 10 (but not 5) days. This was accompanied by increased levels of dopamine in the hippocampus, as well as enhanced dendritic formations (length, number and complexity of synaptic connections) in this same region.
Using the more direct approach of intracranial infusion, Catrine Wernicke and colleagues in Germany found that 9-me-BC has ameliorative effects in a neurotoxin-induced model of Parkinson’s disease, a condition long-associated with insufficient levels of dopamine in specific brain regions such as the striatum.[24] Application of the drug along with the neurotoxin, MPP+, prevented the MPP+-induced death of dopaminergic neurons in both the ventral tegmental area and the substantia nigra. Using an array of biochemical techniques, the authors further suggested that 9-me-BC may work by enhancing mitochondrial function, which is known to be inhibited in patients with Parkinson’s disease. Interestingly, no differences were found between the drug-treated and sham-treated groups, with regards to locomotor activity, as assessed by the chimney test and the tilted plane test.
9-me-BC appears to have multiple mechanisms of action in addition to interacting with the tyrosine hydroxylase pathway of dopamine synthesis. For instance, 9-me-BC is a known inhibitor of monoamine oxidase A,[25] the enzyme responsible for the degradation of the amine neurotransmitters (serotonin, norepinephrine and dopamine). Thus, in addition to enhancing dopamine synthesis, 9-me-BC likely increases circulating levels of dopamine by preventing its breakdown. Further, 9-me-BC appears to decrease toxic inflammation in the brain by decreasing inflammatory cytokines which are known to disrupt cognitive function through microglial accumulation. Thus, 9-me-BC remains an interesting candidate nootropic which has garnered significant attention, though caution is warranted considering a lack of evidence in humans and, as of yet, an unknown efficacy through routes of administration other than injection.
Category 3: A neurogenesis promoter
Falling squarely into the third category of nootropics, drugs which do not directly interact with receptors or enzymes, is NSI-189. NSI-189 is a novel benzylpiperazine-aminopyridine compound, which has been investigated for the treatment of major depressive disorder (MDD) amongst other cognitive deficits. Both in vitro and in vivo data in rodent studies indicate that NSI-189 is capable of stimulating hippocampal neurogenesis, which has been proposed as a potential mechanism for antidepressant drug function.[26] The mechanism by which NSI-189 achieves this remarkable nootropic effect appears to be through stimulation of brain-derived neurotrophic factor (BDNF). Activation of the BDNF pathway is well-known to increase dendritic spine growth and axonal connectivity, both of which are crucial mechanistic underpinnings of learning.
A 2016 phase 1B clinical trial, investigating the efficacy of NSI-189 in a randomized, placebo-controlled study design, indicated significant effects of the drug as orally administered to individuals suffering from MDD either once, twice or three times per day.[27] Outcome measures indicated a marked reduction in MDD symptoms on a self-reported scale. Importantly, using fMRI scanning before and after the treatment, the subjects given NSI-189 also exhibited modestly increased hippocampal volumes. Given the known correlation between MDD, memory dysfunction and depressed neuronal connectivity and plasticity, it is of little surprise that NSI-189 exhibited such positive effects. However, the drug falls into an exceptional category, in that, it can be taken orally, exhibits very few side-effects, and appears to have lasting effects which persist even after the cessation of treatment.
Three other recent animal studies have investigated NSI-189 as a potential pharmacotherapeutic strategy for the treatment of cognitive dysfunction with promising results. First, researchers from the University of South Florida College of Medicine, administered the drug to rats following ischemic stroke. They surmised that enhanced neurogenesis from the drug would be a boon to brain-repair and would therefore accelerate the reversal of cognitive deficits associated with the neurological damage sustained via the stroke.[28] After inducing the stroke through acute occlusion of the middle cerebral artery, the rats were given 30mg/kg NSI-189 orally, beginning 6 hours after the stroke induction and continuing daily for 12 weeks. Rats treated with the drug, as compared to a control group, showed significant improvement after less than one week of treatment, which continued for the remainder of the drug application and, for an additional 12 weeks after withdrawal from the drug, indicating sustained neurological amelioration of stroke-induced cognitive deficits. Specifically, the animals showed improved behavior in the elevated body swing test, forelimb retraction, beam walking and bilateral forepaw grip strength, all common measures of neurological function. These measures were accompanied by enhanced neurogenesis in the hippocampus, in line with previous findings.
In addition to this investigation into stroke-induced cognitive dysfunction, researchers at the University of California, Irvine, tested the efficacy of NSI-189 as a treatment for the cognitive decline associated with radiation therapy.[29] Rats which were exposed to a clinically-relevant level of radiation were tested for mental abnormalities normally occurring following such treatment, such as a reduction in novelty exploration and inhibited learning. As expected, the rats which received the radiation therapy without a remediative drug treatment exhibited decreased levels of novel environment exploration, as well as poor performance in several learning tasks including novel object recognition, object placement, and the temporal order task. Conversely, animals who received NSI-189 orally for 4 weeks, following the radiation exposure, showed levels of novel environment exploration on par with control animals. Similarly, performance in the object recognition and temporal order learning tasks was improved to levels which were not significantly different from the non-irradiated animals, suggesting a full rescue of cognitive function. Mechanistically, NSI-189 also reversed the radiation-induced decrease in neurogenesis, in addition to preventing or reversing neuroinflammation caused by the treatment.
Most recently, researchers in Pomona, California, found significant improvements in cognitive function in a mouse model of Angelman Syndrome, a genetic disorder which, amongst other abnormalities, presents as severe deficits in both motor and cognitive functions.[30] Interestingly, and of particular importance for the potential of NSI-189 as a cognitive enhancer in healthy individuals, the drug was found to enhance long-term potentiation, a form of synaptic plasticity which underlies learning, in the hippocampus of wild-type (control) animals. Angelman Syndrome mice showed similar improvements in hippocampal plasticity in addition to improved function in a fear-conditioning task, indicating that this increased hippocampal function correlates with improved memory. Together with the above investigations, NSI-189 shows remarkable promise as an orally-available promoter of neurogenesis, with a significant ameliorative capability in the cognitive domains of learning, making it a valuable candidate nootropic in addition to a potential therapeutic agent, for a range of disorders.
Conclusions
While there is no dearth of interest in nootropics, there is also a wealth of negative findings regarding putative cognitive enhancers which were eventually found to be of little effect. ADHD drugs such as clonidine and guanfacine, as well as the dopamine agonist Pramipexole, were once touted as potential nootropics, only to be marred later by null findings when tested for cognitive enhancement in healthy individuals.[31] Similarly, countless dietary supplements and vitamins such as omega-3 fatty acids (DHA and EPA), vitamins B6, B9 (folate) and B12, and vitamin E were all regarded with significant interest. Despite some showing nootropic efficacy in animal models or in subjects with cognitive dysfunction (often as a result of dietary deficiencies), these were similarly found to be without effect.[32] This is to say, then, that the business of enhancing cognition in otherwise healthy subjects is rife with oft-unfulfilled promises and that claims of nootropic effect, especially those derived from findings in animal models of disease states or compromised cognition, should be approached with healthy skepticism.
Here, we have presented three potential nootropic agents with variable degrees of evidence and analysis, all of which remain of great interest to those in search of molecules capable of enhancing cognition. Together, Rimonabant and CBD, 9-me-BC, and NSI-189 represent the three major pharmacological classes of nootropic drugs. Each of these appears to have the potential to act as an effective nootropic given the current evidence. While CBD exhibits its most robust effects in a time-dependent manner, others such as NSI-189 potentially imbues positive impacts which extend beyond its dosing period. Both CBD and NSI-189 are orally bioavailable, making them ideal candidates for low-risk nootropic dosing. Conversely, research on 9-me-BC is currently limited to studies using injections of the drug either systemically or directly into the brain. While it still appears to be potentially valuable as a nootropic agent, this detail makes it a less-attractive candidate than some other compounds. Overall, there is a significant amount of interest in the field of nootropics which will continue to drive novel research efforts, hopefully unveiling future results of reliable and significant magnitude for cognitive enhancement.
References
- Froestl, W., Muhs, A., & Pfeifer, A. (2012). Cognitive Enhancers (Nootropics). Part 1: Drugs Interacting with Receptors. Journal of Alzheimer’s Disease, 32(4), 793–887.
- Froestl, W., Muhs, A., & Pfeifer, A. (2014). Cognitive Enhancers (Nootropics). Part 2: Drugs Interacting with Enzymes. Update 2014. Journal of Alzheimer’s Disease, 42(1), 1–68.
- Froestl, W., Pfeifer, A., & Muhs, A. (2014). Cognitive Enhancers (Nootropics). Part 3: Drugs Interacting with Targets other than Receptors or Enzymes. Disease-Modifying Drugs. Update 2014. Journal of Alzheimer’s Disease, 42(4), 1079–1149.
- Terranova, J. P., Michaud, J. C., Le Fur, G., & Soubrié, P. (1995). Inhibition of long-term potentiation in rat hippocampal slices by anandamide and WIN55212-2: reversal by SR141716 A, a selective antagonist of CB1 cannabinoid receptors. Naunyn-Schmiedeberg’s Archives of Pharmacology, 352(5), 576–579.
- Terranova, J. P., Storme, J. J., Lafon, N., Péŕio, A., Rinaldi-Carmona, M., Le Fur, G., & Soubrié, P. (1996). Improvement of memory in rodents by the selective CB1 cannabinoid receptor antagonist, SR 141716. Psychopharmacology, 126(2), 165–172.
- Lichtman, A. H. (2000). SR 141716A enhances spatial memory as assessed in a radial-arm maze task in rats. European Journal of Pharmacology, 404(1–2), 175–179.
- Wolff, M. C., & Leander, J. D. (2003). SR141716A, a cannabinoid CB1 receptor antagonist, improves memory in a delayed radial maze task. European Journal of Pharmacology, 477(3), 213–217.
- Wise, L. E., Iredale, P. A., & Lichtman, A. H. (2008). The cannabinoid CB(1) receptor antagonist CE prolongs spatial memory duration in a rat delayed radial arm maze memory task. European Journal of Pharmacology, 590(1–3), 246–249.
- Takahashi, R. N., Pamplona, F. A., & Fernandes, M. S. (2005). The cannabinoid antagonist SR141716A facilitates memory acquisition and consolidation in the mouse elevated T-maze. Neuroscience Letters, 380(3), 270–275.
- Mazzola, C., Micale, V., & Drago, F. (2003). Amnesia induced by beta-amyloid fragments is counteracted by cannabinoid CB1 receptor blockade. European Journal of Pharmacology, 477(3), 219–225.
- Pisanti, S., Malfitano, A. M., Ciaglia, E., Lamberti, A., Ranieri, R., Cuomo, G., … Bifulco, M. (2017). Cannabidiol: State of the art and new challenges for therapeutic applications. Pharmacology & Therapeutics, 175, 133–150.
- Stern, C. A. J., da Silva, T. R., Raymundi, A. M., de Souza, C. P., Hiroaki-Sato, V. A., Kato, L., … Bertoglio, L. J. (2017). Cannabidiol disrupts the consolidation of specific and generalized fear memories via dorsal hippocampus CB1 and CB2 receptors. Neuropharmacology, 125, 220–230.
- Rossignoli, M. T., Lopes-Aguiar, C., Ruggiero, R. N., Do Val da Silva, R. A., Bueno-Junior, L. S., Kandratavicius, L., … Romcy-Pereira, R. N. (2017). Selective post-training time window for memory consolidation interference of cannabidiol into the prefrontal cortex: Reduced dopaminergic modulation and immediate gene expression in limbic circuits. Neuroscience, 350, 85–93.
- Norris, C., Loureiro, M., Kramar, C., Zunder, J., Renard, J., Rushlow, W., & Laviolette, S. R. (2016). Cannabidiol Modulates Fear Memory Formation Through Interactions with Serotonergic Transmission in the Mesolimbic System. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 41(12), 2839–2850.
- de Carvalho, C. R., & Takahashi, R. N. (2017). Cannabidiol disrupts the reconsolidation of contextual drug-associated memories in Wistar rats. Addiction Biology, 22(3), 742–751.
- Uhernik, A. L., Montoya, Z. T., Balkissoon, C. D., & Smith, J. P. (2018). Learning and memory is modulated by cannabidiol when administered during trace fear-conditioning. Neurobiology of Learning and Memory, 149, 68–76.
- Mori, M. A., Meyer, E., Soares, L. M., Milani, H., Guimarães, F. S., & de Oliveira, R. M. W. (2017). Cannabidiol reduces neuroinflammation and promotes neuroplasticity and functional recovery after brain ischemia. Progress in Neuro-Psychopharmacology and Biological Psychiatry, 75, 94–105.
- Das, R. K., Kamboj, S. K., Ramadas, M., Yogan, K., Gupta, V., Redman, E., … Morgan, C. J. A. (2013). Cannabidiol enhances consolidation of explicit fear extinction in humans. Psychopharmacology, 226(4), 781–792.
- Englund, A., Morrison, P. D., Nottage, J., Hague, D., Kane, F., Bonaccorso, S., … Kapur, S. (2013). Cannabidiol inhibits THC-elicited paranoid symptoms and hippocampal-dependent memory impairment. Journal of Psychopharmacology (Oxford, England), 27(1), 19–27.
- Hamann, J., Wernicke, C., Lehmann, J., Reichmann, H., Rommelspacher, H., & Gille, G. (2008). 9-Methyl-beta-carboline up-regulates the appearance of differentiated dopaminergic neurones in primary mesencephalic culture. Neurochemistry International, 52(4–5), 688–700.
- Goodwill, K. E., Sabatier, C., Marks, C., Raag, R., Fitzpatrick, P. F., & Stevens, R. C. (1997). Crystal structure of tyrosine hydroxylase at 2.3 A and its implications for inherited neurodegenerative diseases. Nature Structural Biology, 4(7), 578–585.
- Sawada, M., Hirata, Y., Arai, H., Iizuka, R., & Nagatsu, T. (1987). Tyrosine hydroxylase, tryptophan hydroxylase, biopterin, and neopterin in the brains of normal controls and patients with senile dementia of Alzheimer type. Journal of Neurochemistry, 48(3), 760–764.
- Gruss, M., Appenroth, D., Flubacher, A., Enzensperger, C., Bock, J., Fleck, C., … Braun, K. (2012). 9-Methyl-β-carboline-induced cognitive enhancement is associated with elevated hippocampal dopamine levels and dendritic and synaptic proliferation. Journal of Neurochemistry, 121(6), 924–931.
- Wernicke, C., Hellmann, J., Ziêba, B., Kuter, K., Ossowska, K., Frenzel, M., … Rommelspacher, H. (2010). 9-Methyl-b-carboline has restorative effects in an animal model of Parkinson’s disease. Pharmacological Reports, 19.
- Keller S, Reichmann H, Gille G. 9-methylb-carboline inhibits monoamine oxidase activity and stimulates the expression of growth factors by astrocytes. Mov. Disord. 25, 619 (2010).
- Malberg, J. E., & Schechter, L. E. (2005). Increasing hippocampal neurogenesis: a novel mechanism for antidepressant drugs. Current Pharmaceutical Design, 11(2), 145–155.
- Fava, M., Johe, K., Ereshefsky, L., Gertsik, L. G., English, B. A., Bilello, J. A., … Freeman, M. P. (2016). A Phase 1B, randomized, double blind, placebo controlled, multiple-dose escalation study of NSI-189 phosphate, a neurogenic compound, in depressed patients. Molecular Psychiatry, 21(10), 1372–1380.
- Tajiri, N., Quach, D. M., Kaneko, Y., Wu, S., Lee, D., Lam, T., … Borlongan, C. V. (2017). NSI‐189, a small molecule with neurogenic properties, exerts behavioral, and neurostructural benefits in stroke rats. Journal of Cellular Physiology, 232(10), 2731–2740.
- Allen, B. D., Acharya, M. M., Lu, C., Giedzinski, E., Chmielewski, N. N., Quach, D., … Limoli, C. L. (2018). Remediation of Radiation-Induced Cognitive Dysfunction through Oral Administration of the Neuroprotective Compound NSI-189, 189(4), 345–353.
- Liu, Y., Johe, K., Sun, J., Hao, X., Wang, Y., Bi, X., & Baudry, M. (2019). Enhancement of synaptic plasticity and reversal of impairments in motor and cognitive functions in a mouse model of Angelman Syndrome by a small neurogenic molecule, NSI-189. Neuropharmacology, 144, 337–344.
- Fond, G., Micoulaud-Franchi, J.-A., Brunel, L., Macgregor, A., Miot, S., Lopez, R., … Repantis, D. (2015). Innovative mechanisms of action for pharmaceutical cognitive enhancement: A systematic review. Psychiatry Research, 229(1–2), 12–20.
- Forbes, S. C., Holroyd-Leduc, J. M., Poulin, M. J., & Hogan, D. B. (2015). Effect of Nutrients, Dietary Supplements and Vitamins on Cognition: a Systematic Review and Meta-Analysis of Randomized Controlled Trials. Canadian Geriatrics Journal: CGJ, 18(4), 231–245.