


Description
The odour-aversion learning apparatus in Drosophila was first developed by Tully and Quinn (Tully and Quinn., 1985) using a combination of a T maze electric-shock reinforcement in a copper wire grid. The MazeEngineers apparatus includes a clear, robust apparatus that includes a holding area and T maze choice areas leading into odor compartments. Two tight vacuum seal lines help create negative pressure for odor diffusion. The copper training tube can be easily electrified using a simply included plug. Included in your order is a clamp to ensure minimal odor release in the testing phase.
Additional paradigms for choice testing only and Light Dark paradigms are also available.
Odors not included. Olfactometers upon request.
A: Training Tube: Inner surface covered with an electrifiable copper grid
B: T Maze Choice Point for testing relative odor preferences
C: Sliding Center Compartment: For transfering flies from the training tube to the choice point and odor tube
D: Odor tubes: odor cups placed here
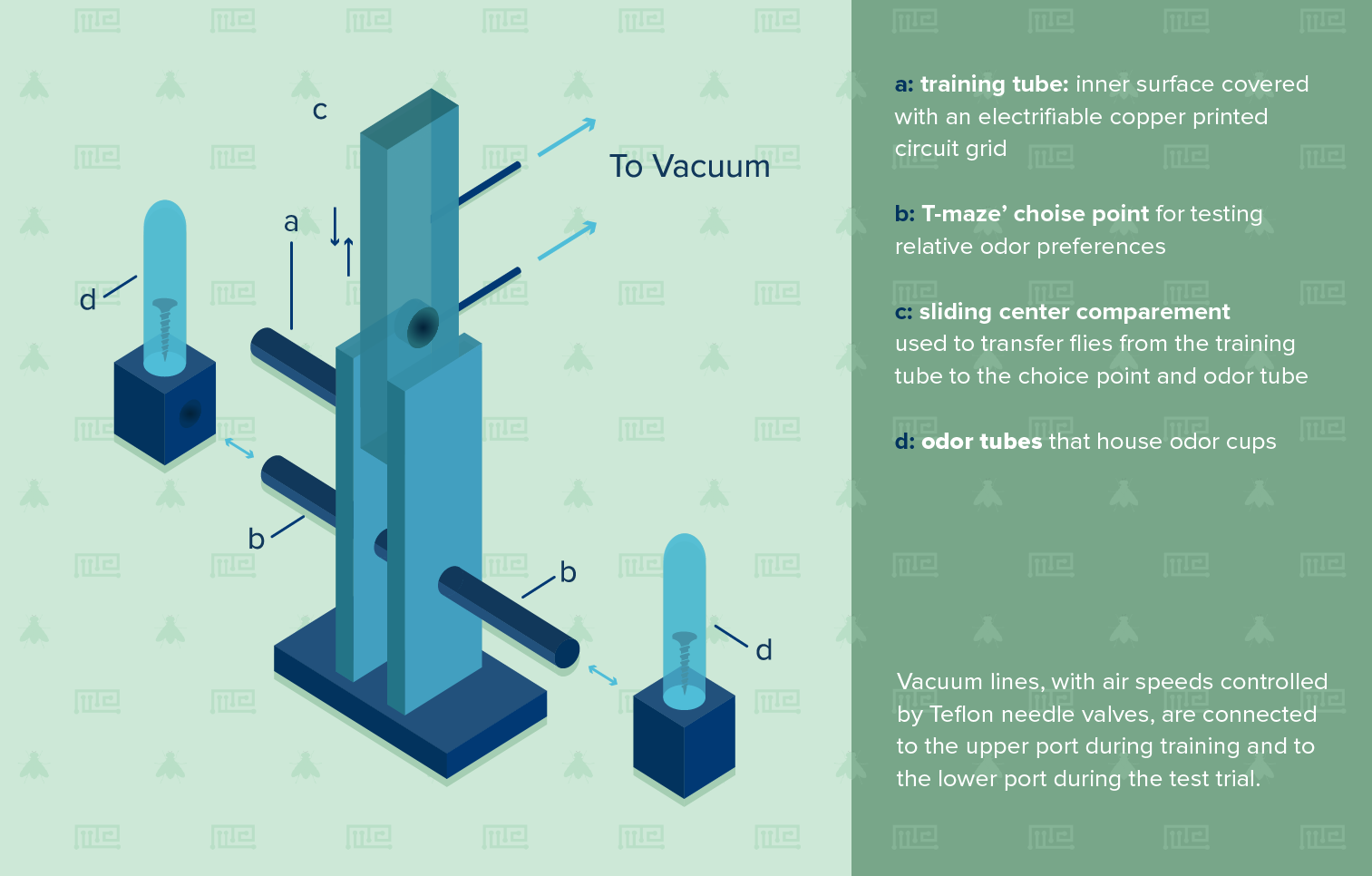
Note: Vacuum lines are connected to the upper port (on the sliding doors) during training and the the lower port during trial .
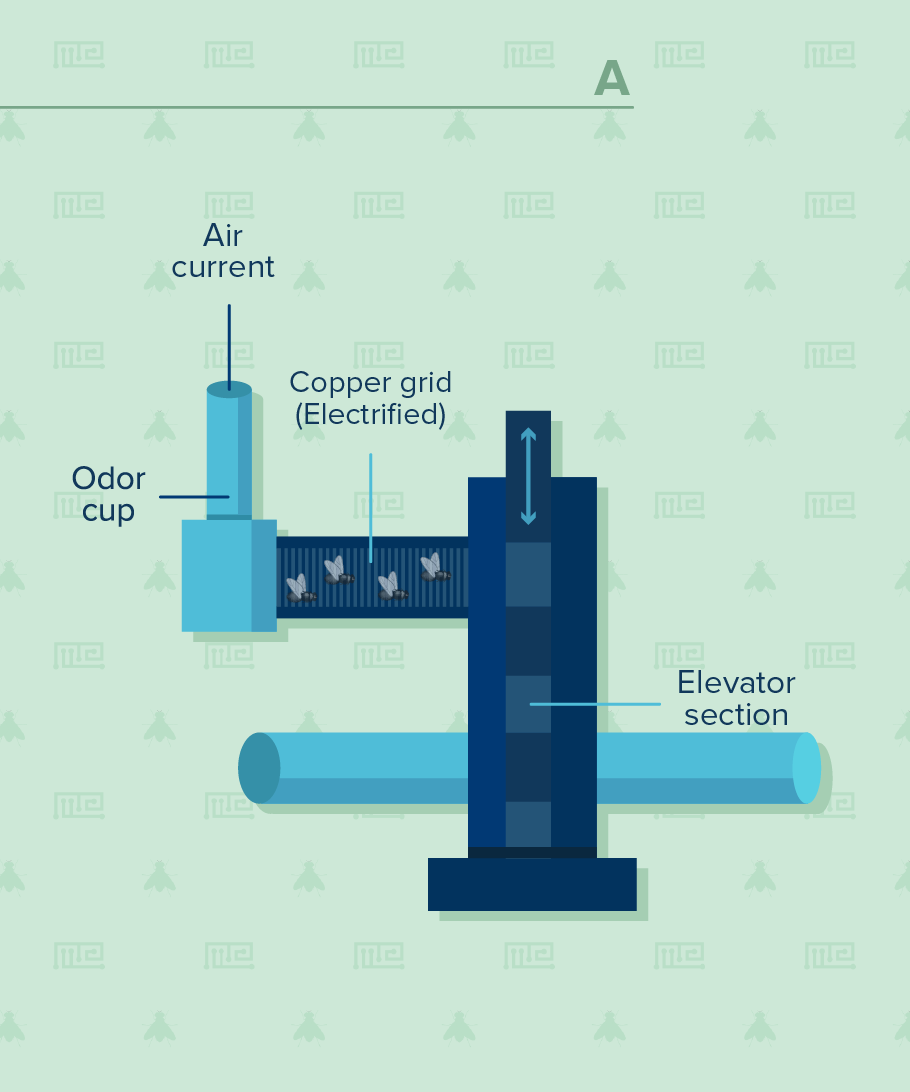
Training Phase
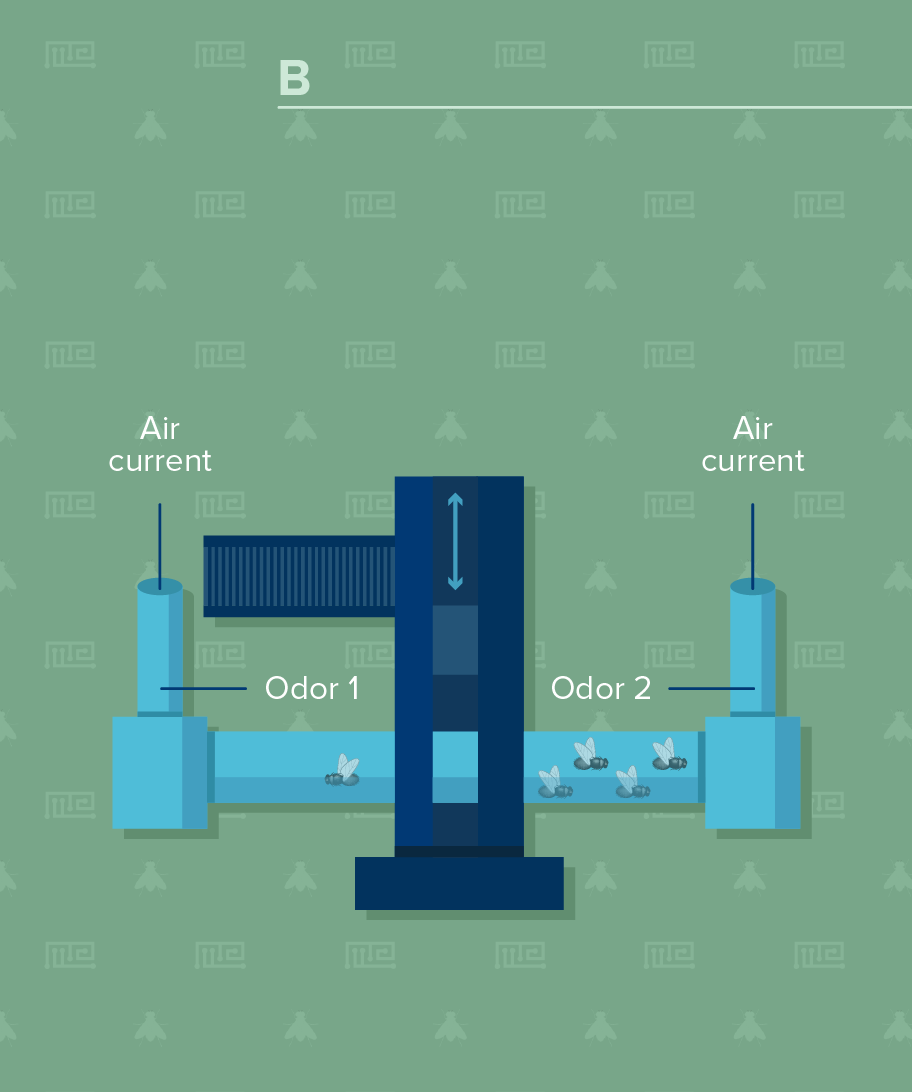
Testing Phase
Price & Dimensions
Complete system
$ 4900
Per Month- 2 Odor chambers
- T maze choice area
- Vacuum and tubing (0.6-6L/min)
- Secure clamp
- Copper training tube
- Control Box
- Software
T maze Choice Area only
$ 1499
Per Month- 2 Odor chambers
- T maze choice area
- Vacuum and tubing (0.6- 6L/min)
- Secure clamp
Light Dark T maze Choice
$ 1599
Per Month- 2 Odor chambers
- T maze choice area
- Manual light switch for 1 chamber
- Vacuum and tubing (0.6-6L/min)
- Secure clamp
Documentation
Introduction
Olfactory aversive conditioning in Drosophila is based on the work of Tully and Quinn (Tully and Quinn., 1985). The protocol typically involves a binary T-maze choice. In conditioning experiments, flies learn to associate one of two odors with an electrical shock in a training tube.
Assessments of learning and of short-, mid-and long-term memory are possible. Evaluation of learning is normally performed immediately after training; determination or short-term memory is done 30 min-1 hr after training, and evaluation of mid-term memory is performed at 3 hr post-training. Moreover, the labile phase of memory as a result of a one-cycle training can be detected for up to 7 hr. In addition, long-term memory lasting up to a week can be formed by repetitive training cycles with gaps between them. In the alternative, training without gaps leads to the formation of anaesthesia-resistant memory, which is similar to long-term memory.
The test should be performed under dim-red light or darkness to enable visualization of the flies, but prevents them from seeing the experimenter. The test requires opaque open cylindrical tubes connected to a T-maze. An air pump should be attached to the tubing to ensure the diffusion of the odors. A proper airtight seal is therefore critical. An electrifiable grid is used to induce the shocks, whose onset, intensity and duration are controlled by the investigator.
The experimental protocol entails a training period starting with adaptation to the tube and the airflow. Exposure to the first odor and the electrical shock is followed by the presentation of the second odor associated with no shock. In case learning is being assessed, a brief rest period precedes the testing phase. At this point, the flies choose one of two tubes containing either odor used in testing. If assessing memory, flies should be kept in the dark at a lower temperature until testing.
Overall, Drosophila is a widely used model in neurobehavioral assessments as it enables genetic, behavioral, dietary and pharmacological manipulations. Furthermore, Drosophila presents other advantages, such as inexpensive maintenance of large sample sizes, small size and rapid life cycle that enables longitudinal studies, and the ability to study discrete neural pathways and structures (Buchanan et al., 2015).
Apparatus and Equipment
Analysis of operant conditioning in Drosophila typically requires opaque open cylindrical tubes connected to a T-maze. An airtight seal should be ensured inside compartments. The T-maze should be attached to an vacuum via tubing to enable exposure to the odors. An electrifiable grid is used to induce the adverse stimuli. Malik and Hodge (2014) suggest placing copper grids inside the training tube. These grids should be attached to a switch-box connected to an electric stimulator via crocodile clips. The experimenter should have control over shock presentation, intensity and duration. A voltmeter can be used to ensure that the apparatus is delivering the required shock (Malik and Hodge, 2014).
Training Protocol
For studies using transgene expression or heat-activated ion channels to stimulate neurons, the flies should be reared at 18-23 °C to prevent the expression of transgenes or maintain the channels in an inactive status, respectively; testing should be done at 30-33 ºC (Malik and Hodge, 2014, Buchanan et al., 2015). Similar considerations are in place if using the Shibiretemperature sensitive phenotype to block synaptic output (Malik and Hodge, 2014, Buchanan et al., 2015). If not testing immediately after training, Malik and Hodge (2014) recommend storing the flies at 25 ºC and 70% relative humidity with 12:12 hr light:dark conditions. Experiments should be performed under dim-red light or darkness to enable observation of the flies, but preventing them from seeing the investigator (Pitman et al., 2009, Malik and Hodge, 2014). This protocol enables simultaneous testing of 100 flies (Pitman et al., 2009).
Training
Training in Drosophila aversive conditioning starts with introducing the flies into the training tube, which is attached to the T-maze. The flies should be allowed a minimum of 90 sec to adapt to the tube and the airflow (Malik and Hodge, 2014) before initial simultaneous exposure to odor 1 (+) and an electric shock. Pitman et al. (2009) recommend 1 min exposure to odor 1 with twelve 1 sec electric shocks (5 sec intervals). Malik and Hodge (2014) recommend using 60-V shocks consisting of twelve 1.25 sec pulses with 3.75 sec inter-pulse intervals. Subsequently, the flies are exposed to 30 sec of fresh air, followed by another 1 min exposure to odor 2 (-) without the electric shock. The flies should then be allowed an additional 30 sec rest period before testing.
If assessing performance index (see Sample data) within a group of flies, a separate population is trained with the + and − odors reversed and a final performance index will be the average of the two reciprocal half experiments (Pitman et al., 2009).
Common odor choices include 4-methylcyclohexanol, 3-octanol and quinine hydrochloride (Pitman et al., 2009, van Swinderen, 2011).
Testing
In testing, Pitman (2009) recommend transporting the flies in the elevator to the T-maze where they are allowed 2 min to choose between tubes containing either of the two odorants used during conditioning. Conversely, Malik and Hodge (2014) recommend moving the flies from the training chamber into the central chamber of the T-maze by turning the T-maze on its side and gently banging the bottom of the apparatus on a soft surface. As in training, the flies should be maintained in the central chamber for 90 sec. To measure learning, the investigator should move the flies to the choice point of the T-maze, to enable simultaneous exposure to both odors. The test period – evaluation of which side the flies choose – should be conducted for 2 min. The flies should then be collected in each arm of the T-maze and in the central compartment into food vials and counted.
To measure memory, flies should be transferred to food vials without yeast and stored in the dark until testing. If assessing long-term memory, Malik and Hodge (2014) recommend administering 5 cycles of training with or without a 15 min inter-cycle interval. The flies should be kept at 18 °C and 70% humidity in the dark. Prior to testing, the investigator should move the flies back to 25 °C and allow them to acclimatize for at least 1 hr.
After testing, Malik and Hodge (2014) recommend cleaning odor cups with hot water and odorless detergent. After drying, the investigator should coat the cups with sigmacote. In addition, T-maze tubes and odor blocks should be occasionally cleaned with hot water and odorless detergent as well.
Modifications
Instead of aversive stimuli, sucrose reward in food-deprived flies can be used (Tempel et al., 1983). Training and testing protocols are similar to aversive conditioning. Compared to shocks, sucrose leads to longer memory retention (Tempel et al., 1983, Malik and Hodge, 2014). The work of Colomb et al (2009) showed that both sugar responsiveness and short-term memory increase with starvation length before conditioning. Furthermore, the authors observed that the protein dependency of long-term memory is independent of repetition and spacing of training sessions, on the starvation duration and on the strength of the unconditioned stimulus.
Sucrose reward can also be used to assess the proboscis extension reflex, based on the observation that flies extend their proboscis when sugar is presented to gustatory receptors on the foreleg tarsi. This extension depends on sugar concentration and the concentration of a bitter substance added to the sugar. Flies are starved and immobilized on slides. This paradigm can also be associated with odors as above (Wang et al., 2004, Chabaud et al., 2006).
In alternative to shocks, analysis of operant conditioning in Drosophila can be done with light. In this regard, van Swinderen (2011) reported a paradigm called aversive phototaxic suppression, which uses light/dark association to assess visual perception. The flies learn to associate a lit chamber with an aversive stimulus (quinine hydrochloride), which is present throughout the study. The fly’s ability to avoid phototaxis is evaluated by the number of dark/light choices.
Sample Data
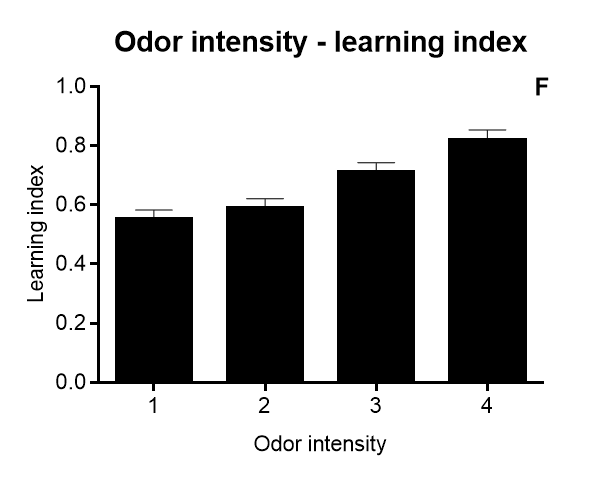
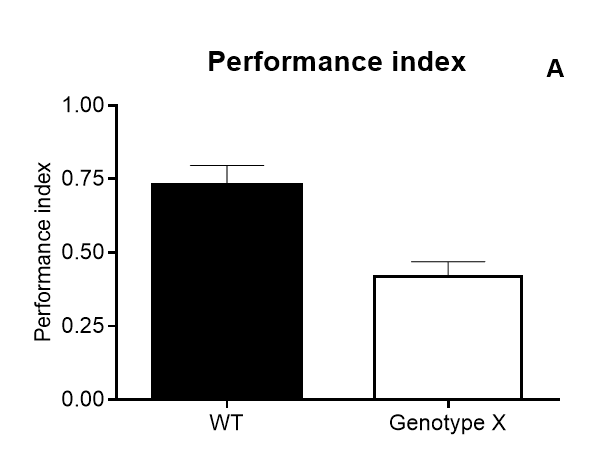
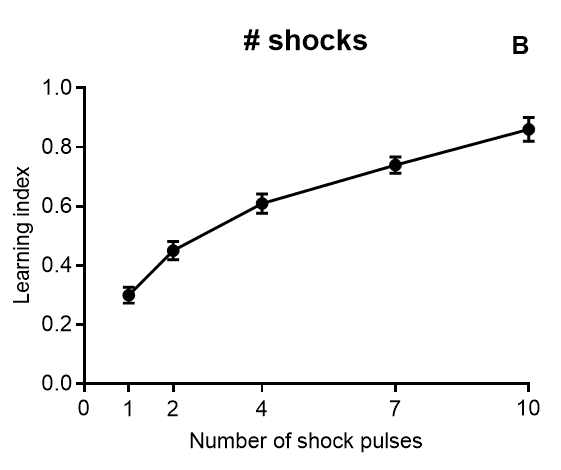
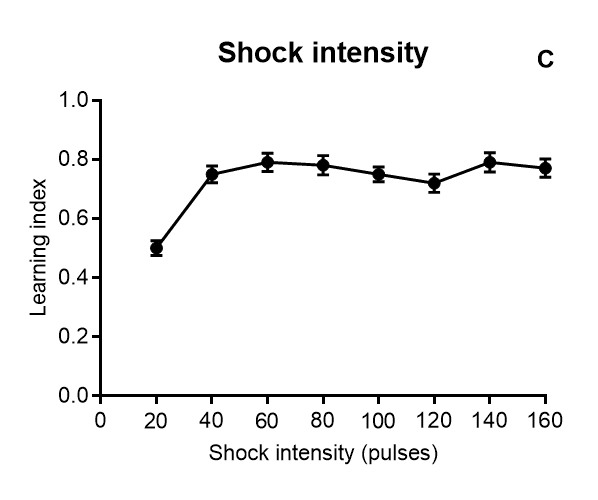
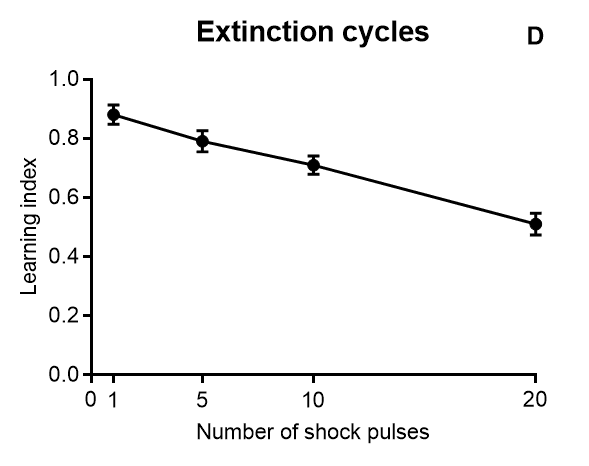
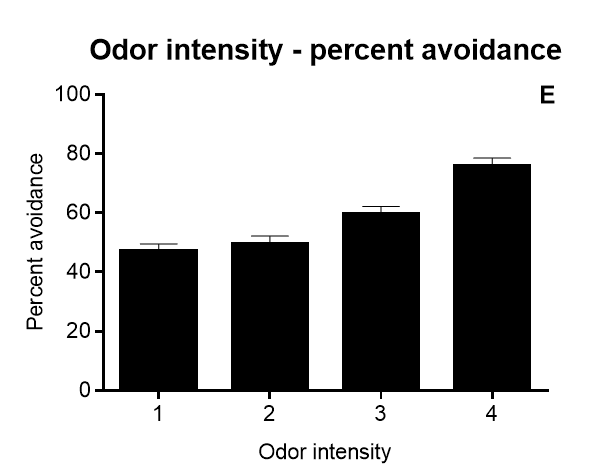
Results of Drosophila’s olfactory aversive conditioning can be presented in diverse ways, depending on the parameters under study. The protocol can be used to compare learning in different genotypes, which would be plotted as a performance index taking into account preference and avoidance of either odor (A) (Malik and Hodge, 2014). Performance index of 1 represents learning in every fly. In alternative, assessment of learning can be shown as a function of the number of shocks (B) and shock intensity (C) (Tully and Quinn, 1985). Furthermore, the investigator can analyze extinction by performing various numbers of extinction cycles (which lack the shock) after the training cycle (D), and/or measure learning as a function of odor intensity (E-F) (Tully and Quinn, 1985). Results are shown as mean ± standard error of the mean:
Genotypic differences
Careful control for genetic background should to be done before comparing flies from different origins. In this regard, the study of Tully and Quinn (1985) showed that, compared to wild-type (C-S) Drosophila, mutant strains dunce, rutabaga and amnesiac have faster memory retention decay, particularly when assessing learning, short-, and mid-term memory. Furthermore, the researchers compared learning scores of 15 mutant strains to wild-type Drosophila. Of these, Berlin strain performed the worst. Furthermore, flies homozygous for the turnip learning mutation learn about as well as rutabaga flies, while tur/C-S heterozygotes do not show learning differences when compared to C-S flies. Conversely, the double learning mutant dnc TM 1, rut, performs worse than either single mutant.
In a study using sugar (positive) conditioning to assess memory deficits of mutant flies, Colomb et al. (2009) showed that radish mutants have abnormal sugar responsiveness. In addition, crammer and tequila mutants, which had previously been shown to be deficient for aversive long-term memory, presented defects in sugar responsiveness and in short-term memory.
Strengths & Limitations
Strengths
Operant conditioning in Drosphila is a simple and high-throughput method of studying learning memory processes. The ability to use aversive and positive (sucrose) stimuli associated with the odors further enables flexibility of the experimental design.
Overall, the use of Drosophila enables reliable experimental designs, which are potentiated by easy and inexpensive generation and maintenance of large experimental groups. Drosophila’s rapid life cycle facilitates the design of time-efficient longitudinal studies, including the study of neurodegenerative diseases.
Limitations
Although the specific role of discrete neural structures can be disentangled in neurobehavioral assessments in Drosophila, the lack of clear homology to the human brain is a limitation to the use of this model.
Summary and Key Points
- Operant conditioning in Drosophila is a popular and reliable method to assess learning and memory. The study of neurodegenerative processes is made possible due to the fly’s rapid life cycle. Longitudinal studies, including genetic, behavioral, dietary and pharmacological manipulations are a further possibility in using this model.
- The test is performed under dim-red light or darkness using opaque, open and airtight sealed cylindrical tubes connected to a T-maze. An air pump enables exposure of the odors, whereas an electrifiable grid is used to induce the shocks, which are controlled by the investigator.
- The protocol included a training period starting with adaptation to the tube and the airflow. Exposure to the first odor associated with the electrical shock is followed by the second odor without shock. Assessment of learning and variable types of memory (short-, mid- or long-term) can be performed. During testing, the flies choose one of two tubes containing either odor used in testing.
- Results presented when using operant conditioning in Drosophila can include analysis of a performance index taking into account preference and avoidance of either odor. Furthermore, data regarding learning ability can be presented as a function of the number of shocks, their intensity, or odor insensity. Analysis of extinction (after a set number of training cycles lacking the shock) is also possible.