
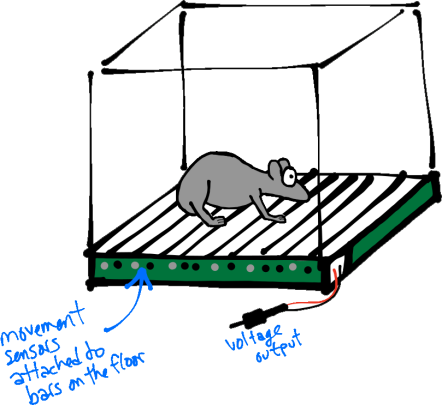
A startle chamber. Sensors connected to the floor bars detect movement and output voltage. Typically, footshocks (see below) can also delivered to the floor bars, although the mechanism for doing so is not depicted here.
Today’s topic is the acoustic startle reflex response (in mice/rats). Many of you may be familiar with this term, but may not really know what it is or why it continues to be a topic of interest in neuroscience research. Let’s start with the basics.
What is it?
The acoustic startle reflex (ASR) is actually pretty simple. When mice/rats are surprised by a loud noise (Fig. 1, left; Meow!), their facial and skeletal muscles rapidly contract. This includes closing of the eyes, stiffening of the neck and body (Fig. 1, left; startled mouse), and other physical changes that might protect the animal and help prepare it for a fight/flight response.
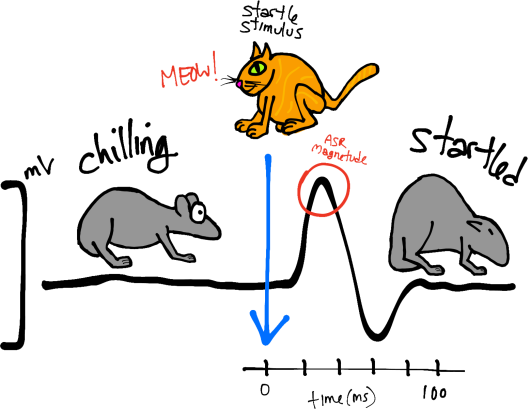
Figure 1. Left, schematic of the startle response, including the quantified response (black line = voltage trace) and characteristic behavior.
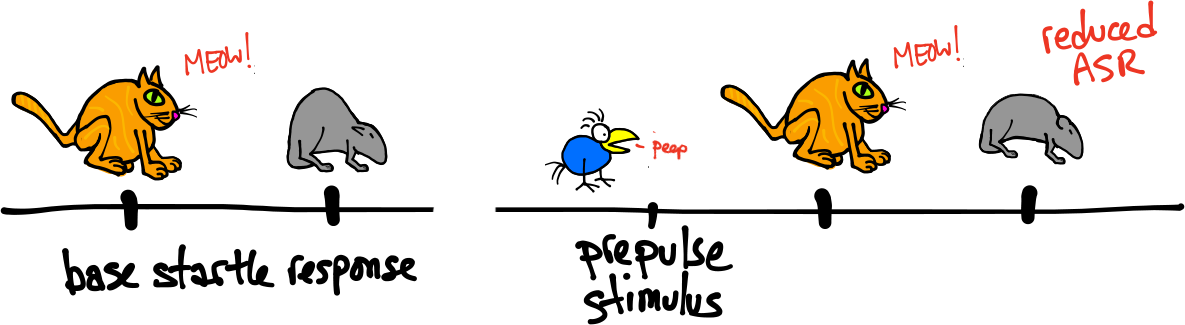
Pre-pulse inhibition. Left, baseline ASR. Right, reduced ASR following a pre-pulse
How is it measured?
Typically, ASRs are characterized by their magnitudes (and latencies). The magnitude of a whole-body movement can be measured by placing animals in startle chambers (Fig. 1, right) rigged with sensors (piezoelectric transducers) that convert small movements to voltage. Thus, the magnitude of the change in voltage represents the size of the ASR (Fig. 1, left; peak of the black trace).
What is the basic neural circuitry?
Primary circuitry can be summarized in this figure adapted from Koch (1999) (blue arrows indicate the fastest route from stimulus to response
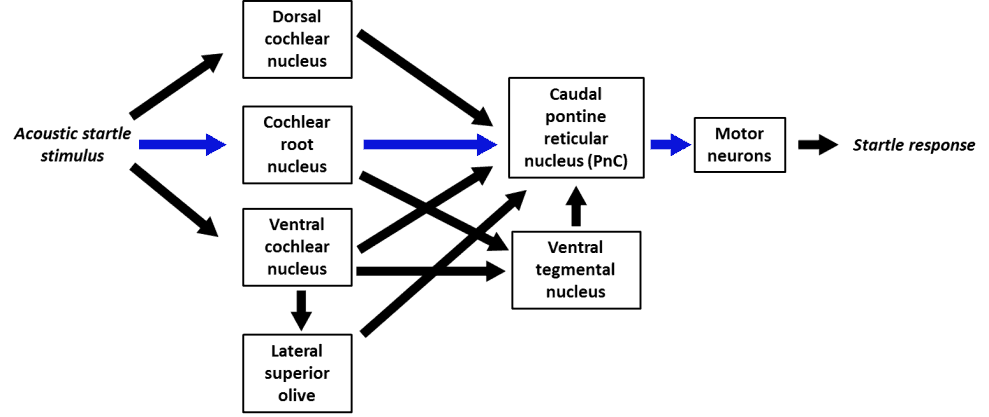
Primary circuitry, adapted from Koch (1999) (blue arrows indicate the fastest route from stimulus to response
Beyond the basics:
Ok, now let’s move on to the not-so-basics. What makes the ASR interesting and useful is that, while the characteristics of the response are almost always the same, a given startling sound can elicit ASRs that vary in magnitude, depending on the situational context (e.g. the animal’s mood or the presence of additional sensory stimuli). Because of this, it has become a popular behavioral tool for studying sensorimotor response (neuro)plasticity. Doing so usually involves using standard manipulations that either attenuate or enhance the ASR.
Sensitization:
Delivering a fearful stimulus (usually footshocks, not blue monsters) after an ASR causes subsequent responses to the startle stimulus (usually a loud tone, not cute orange cats) to be greater in magnitude (sensitized). Sensitization lasts for a while after the fearful stimulus is no longer present. Lesions and drug studies show that sensitization of the ASR does not occur when the amygdala does not function properly.

Sensitization
Fear Potentiation:
Pairing the startle stimulus with a fear-conditioned stimulus (CS, usually a light paired with footshock, see Fig. 4, left) will induce a startle response that is greater than what occurs with the startle stimulus alone. Different from sensitization, this enhancement does not last long after the conditioned stimulus is gone. The greatest effect occurs when the timing between the CS and the startle stimulus matches that of the original pairing between the CS and the aversive stimulus. The amygdala is required both for the association between the CS and the aversive stimulus, as well as for the potentiated ASR after successful fear conditioning. Output from the amygdala projects to the ASR circuit at the level of the pontine reticular nucleus.

Enhanced ASRs occur when a startle stimulus is paired with a fear-conditioned stimulus.
Habituation:
Repetition of the startle stimulus eventually leads to a weaker ASR. The literature distinguishes between 2 types of habituation: Short-term and long-term. Short-term habituation occurs within a single session, while long-term habituation occurs between sessions. Habituation, like sensitization, does not require any learned associations.

Reduced ASR after stimulus repetition. Here a single session is depicted, so this is shirt-term habituation.
Pre-pulse Inhibition:
Delivering a weak sensory stimulus before the startle stimulus reduces the subsequent ASR. The pre-pulse stimulus is usually a low decibel tone, but any stimulus that does not induce a startle response is ok. The time between the pre-pulse stimulus and the startle stimulus can range between approximately 6 ms and 2 seconds.
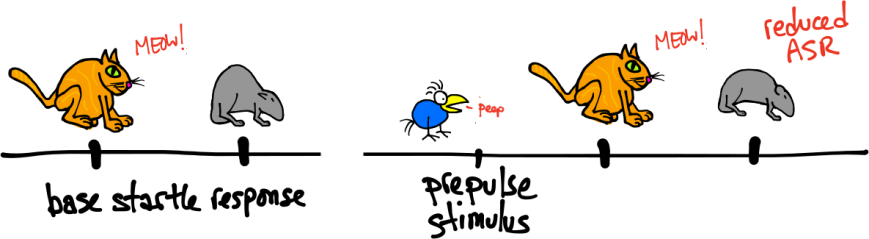
Pre-pulse inhibition. Left, baseline ASR. Right, reduced ASR following a pre-pulse
Take-home message:
The startle response is a useful tool for studying stimulus-response neural plasticity. The response circuit is well characterized, and paradigms that modulate the response magnitude are used in conjunction with lesions and pharmacological manipulations to determine which brain regions regulate the response in different contexts, as well as what the relevant receptors and neural transmitters are. Answering these questions may help understand certain human disorders such as schizophrenia that show characteristic abnormalities in startle responses. Future blog posts will cover these findings in more detail (stay tuned!).
References/Further reading:
Koch M (1999). The neurobiology of startle. Prog Neurobiol. 59:107-28.
Koch M, Schnitzler HU (1997). The acoustic startle response in rats–circuits mediating evocation, inhibition and potentiation. Behav Brain Res. 89:35-49.

Koch, M. (1999). The neurobiology of startle Progress in Neurobiology, 59 (2), 107-128 DOI: 10.1016/S0301-0082(98)00098-7